Volume 7, Number 1—February 2001
Synopsis
Geographic Subdivision of the Range of the Malaria Parasite, Plasmodium vivax
Figure 2
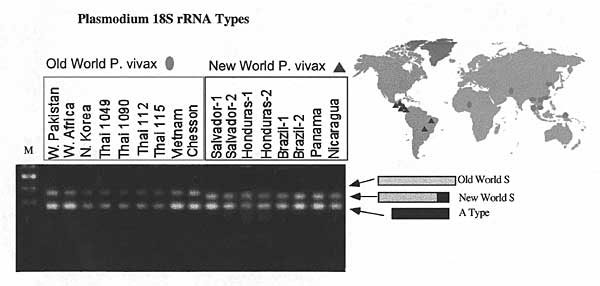
Figure 2. Sequences of Plasmodium vivax isolates are distinguished by variation in the 3' end of the S-type rRNA gene (10). The S-type gene is longer in Old World isolates and in P. simium. Oligonucleotide #902 (5'CAGCAAGCTGAATCGTAATTTTAA3') was used to detect type A rRNA, and #743 (5'ATCCAGATCCAATCCGACATA3') and #901 (5'GATAAGCACAAAATAGCGAAATGC3') were used to differentiate the two S-type rRNAs in membrane blot hybridization. American Type Culture Collection reference numbers not designated in the Figure 1 legend are as follows: Thai R112, Thai R115, Honduras-1 T09794, Honduras-2 T10595, Brazil-1 T40695, and Thai K1090. Haiti, Brazil-2, and the West African isolates came directly from the Centers for Disease Control and Prevention (CDC).*† *The P. vivax SAL-1 strain, Africa strain, Pakistan strain, Brazil strain, and New World monkey vivax, P. simium, were from CDC. P. vivax Vietnam strain, Chesson strain, and Panama strain were from the American Type Culture Collection. The four Thai strains were from Walter Reed Medical Center; they were collected from four geographically separate locations in Thailand and their immunologic characteristics described with regard to the circumsporozoite protein gene (9). The heparinized blood samples were stored in 5% glycerolyte at -70°C. †Purified DNA from frozen blood samples was processed with DNAzol reagents (GIBCO-Bethesda Research Laboratory, Gaithersburg, MD) according to the manufacturer's instructions. The partial sequence of 18S rRNA genes that covers variable regions 7 and 8 was amplified with a pair of genus-conserved primers, #841 and #844 (10). The sequence of an open reading frame (ORF 470) on the 35-kb plastid-like DNA was amplified with a pair of oligonucleotide primers #1274 (5' GTAAAATTATATAAACCACC 3') and #1273 (5' GCACAATTTGAACGTAC 3') conserved in plastid-like organelle in Apicomplexa (11). The sequence of circumsporozoite protein genes was amplified with oligonucleotide primers #1157 (5'AATGGAGTAAACTTCAATAATGTA 3') and #1160 (5' CTCCACAGGTTACACTGCATG 3'). The reaction was set up in a 100-L reaction volume containing 20-50 ng DNA, 200 M of each deoxynucleoside triphosphate, 50 mM KCl, 10 mM Tris-HCl (pH8.3), 2 mM MgCl2, and 2.5 U Taq DNA polymerase (Perkin-Elmer Cetus, Norwalk, CT) at a three-step cycling with the following parameters: 94C/1 min for denaturation, 50C/1 min for annealing, and 72C/1-2 min for elongation for a total of 30 cycles. Conditions for separation of polymerase chain reaction products on agarose electrophoresis and 32P-ending labeled probe hybridization were described (8,10). Oligonucleotide #902 (5'CAGCAAGCTGAATCGTAATTTTAA3') was used to detect type-A rRNA, and #743 (5'ATCCAGATCCAATCCGACATA3') and #901 (5'GATAAGCACAAAATAGCGAAATGC3') were used to differentiate the two S-type rRNAs in membrane blot hybridization. Oligonucleotide #1163 (5'AGAGCAGCTGGACAGCCAGCA3') and #1165 (5'GCWGGCAATCAACCAGGAGCA3') were used as probes to differentiate Sal-1 (classic) and PVK247 (variant) types of circumsporozoite protein genes, respectively. The gene coding for 18S ribosomal RNA was amplified from the DNA of Thai isolates with oligonucleotides #566, 5'GGA TAA CTA CGG AAA AGC TGT AGC3', and #570, 5'CGA CTT CTC CTT CCT TTA AAA GAT AGG3', as the 5' and 3' end primers, respectively. Both primers are conserved for the genus Plasmodium and cover most of the transcribed sequences from approximately 140 downstream of the 5' end to 40 bp upstream of the 3' end.
References
- Bruce-Chwatt LJ. Essential malariology. New York: John Wiley and Sons; 1985.
- Mayr E. Animal species and evolution. Cambridge: Belknap Press of Harvard University Press; 1963.
- Killick-Kendrick R. In: Peters W, editor. Rodent malaria. London: Academic Press; 1978.
- Collins WE, Skinner JC, Pappaioanou M, Ma NS, Broderson JR, Sutton BB, Infection of Aotus vociferans (karyotype V) monkeys with different strains of Plasmodium vivax. J Parasitol. 1987;73:536–40. DOIPubMedGoogle Scholar
- Collins WE, Warren M, Huong AY, Skinner JC, Sutton BB, Stanfill PS. Studies of comparative infectivity of fifteen strains of Plasmodium vivax to laboratory-reared anopheline mosquitoes, with special reference to Anopheles culicifacies. J Parasitol. 1986;72:521–4. DOIPubMedGoogle Scholar
- Collins WE, McClure H, Strobert E, Skinner JC, Richardson BB, Roberts, et al. Experimental infection of Anopheles gambiae s.s., Anopheles freeborni, and Anopheles stephensi with Plasmodium malariae and Plasmodium brasilianum. J Am Mosq Control Assoc. 1993;9:68–71.PubMedGoogle Scholar
- Li J, Wirtz RA, McCutchan TF. Analysis of malaria parasite RNA from decade-old giemsa-stained blood smears and dried mosquitoes. Am J Trop Med Hyg. 1997;57:727–31.PubMedGoogle Scholar
- Li J, Wirtz RA, McConkey GA, Sattabongkot J, McCutchan TF. Transition of Plasmodium vivax ribosome types corresponds to sporozoite differentiation in the mosquito. Mol Biochem Parasitol. 1994;65:283–9. DOIPubMedGoogle Scholar
- Wirtz RA, Burkot TR, Andre RG, Rosenberg R, Collins WE, Roberts DR. Identification of Plasmodium vivax sporozoites in mosquitoes using an enzyme-linked immunosorbent assay. Am J Trop Med Hyg. 1985;34:1048–54.PubMedGoogle Scholar
- Li J, Wirtz RA, McConkey GA, Sattabongkot J, Waters AP, Rogers MJ, Plasmodium: genus-conserved primers for species identification and quantitation. Exp Parasitol. 1995;81:182–90. DOIPubMedGoogle Scholar
- Wilson RJM, Denny PW, Preiser PR, Rangachari K, Roberts K, Roy A, Complete gene map of the plastid-like DNA of the malaria parasite Plasmodium falciparum. J Mol Biol. 1996;261:155–72. DOIPubMedGoogle Scholar
- Waters AP, McCutchan TF. Partial sequence of the asexually expressed SU rRNA gene of Plasmodium vivax [published erratum appears in Nucleic Acids Res 1989 May 11;17:3630-1]. Nucleic Acids Res. 1989;17:2135. DOIPubMedGoogle Scholar
- Escalante A, Barrio E, Ayala FJ. Evolutionary origin of human and primate malarias: evidence from the circumsporozoite protein gene. Mol Biol Evol. 1995;12:616–26.PubMedGoogle Scholar
- Vaidya AB, Morrisey J, Plowe CV, Kaslow DC, Wellems TE. Unidirectional dominance of cytoplasmic inheritance in two genetic crosses of Plasmodium falciparum. Mol Cell Biol. 1993;13:7349–57.PubMedGoogle Scholar
- Gupta S, Ferguson N, Anderson R. Chaos, persistence and evolution of strain structure in antigenically diverse infectious agents. Science. 1998;280:912–5. DOIPubMedGoogle Scholar
- Lal AA, de la Cruz VF, Collins WE, Campbell GH, Procell PM, McCutchan TF. Circumsporozoite protein gene from Plasmodium brasilianum. Animal reservoirs for human malaria parasites? J Biol Chem. 1988;263:5495–8.PubMedGoogle Scholar
- Escalante AA, Freeland DE, Collins WE, Lal AA. The evolution of primate malaria parasites based on the gene encoding cytochrome-b from the linear mitochondrial genome. Proc Natl Acad Sci U S A. 1998;95:8124–9. DOIPubMedGoogle Scholar
- Kain KC, Brown AE, Webster HK, Wirtz A, Keystone JS, Rodriguez MH, Circumsporozoite genotyping of global isolates of Plasmodium vivax from dried blood specimens. J Clin Microbiol. 1992;30:1863–6.PubMedGoogle Scholar
- Simpson GC. Tempo and mode in evolution. New York: Columbia University Press; 1944.
- Coatney GR, Collins WE, Warren M, Contacos PG. The primate malarias. Bethesda: Dept of Health, Education and Welfare (US); 1971.
- Gonzalez-Ceron L, Rodriquez MH, Nettel JC, Villarreal C, Kain KC, Hernandez JE. Differential susceptibilities of Anopheles albimanus and An. pseudopunctipennis to infections with coindigenous Plasmodium vivax variants VK210 and VK247 in southern Mexico. Infect Immun. 1999;67:410–2.PubMedGoogle Scholar
¹The biologic diversity inherent in P. vivax already justifies the use of a trinomial system for naming its members that includes the designation of subspecies, a taxonomic character given formal recognition in the International Rules of Zoological Nomenclature. A subspecies is a population or group of populations inhabiting a geographic subdivision of the range of a species and differing from other populations by diagnostic morphologic characteristics.
²The designation of separate species does not require that the two organisms cannot mate and produce viable progeny, only that this does not happen with frequency in natural situations.