Volume 13, Number 7—July 2007
Synopsis
Thottapalayam Virus, a Prototype Shrewborne Hantavirus
Cite This Article
Citation for Media
Abstract
Thottapalayam virus (TPMV) has been placed in the genus Hantavirus of the family Bunyaviridae by virtue of its morphologic features and overall genetic similarities to well-characterized rodentborne hantaviruses. This virus has been isolated from the Asian house shrew (Suncus murinus); however, whether TPMV is naturally harbored by an insectivore host or represents spillover from a rodent reservoir host is unknown. Our analysis of published and unpublished data on the experimental host range, genetics, and molecular phylogeny of TPMV supports coevolution of TPMV with its nonrodent reservoir host. Future studies on the epizootiology of TPMV and investigations of new shrewborne hantaviruses will provide additional insights into the evolutionary origin of hantaviruses in their rodent and insectivore reservoir hosts. Such investigations may also provide clues about determinants of hantavirus pathogenicity and virulence.
Viruses in the genus Hantavirus, similar to other members of the family Bunyaviridae, have a negative-sense, single-stranded RNA genome in 3 segments designated large (L), medium (M), and small (S), which encode an RNA-dependent RNA polymerase, envelope glycoproteins (Gn, Gc) and nucleocapsid (N) protein, respectively (1,2). Each viral genomic segment has the identical 3′-terminal sequence of AUCAUCAUCUG, which is unique to hantaviruses (3). However, unlike the >200 other members in this virus family, most of which have arthropod vectors, each genetically distinct hantavirus is harbored by 1 or a few closely related rodent species with which it appears to have coevolved (4,5). Hantaan virus (HTNV) shares a multimillennium relationship with the striped field mouse (Apodemus agrarius), Dobrava virus (DOBV) with the yellow-necked field mouse (Apodemus flavicollis), Seoul virus (SEOV) with the Norway rat (Rattus norvegicus), Thailand virus (THAIV) with the bandicoot rat (Bandicota indica), Puumala virus (PUUV) with the bank vole (Myodes glareolus, formerly Clethrionomys glareolus), Tula virus (TULV) with the European common vole (Microtus arvalis), Prospect Hill virus (PHV) with the meadow vole (M. pennsylvanicus), and Sin Nombre virus (SNV) with the deer mouse (Peromyscus maniculatus).
Many other rodent-hantavirus associations are known, including the recent discovery of a hantavirus in the African wood mouse (Hylomyscus simus) (6). Until recently, the 1 exception that did not have a confirmed rodent association has been Thottapalayam virus (TPMV), which was isolated from an Asian house shrew or musk shrew (Suncus murinus) captured in 1964 during a survey for Japanese encephalitis virus in southern India (7). TPMV has been classified as a hantavirus by virtue of its ultrastructural features (8) and overall genetic similarities with well-characterized rodentborne hantaviruses (9,10). Although isolation of TPMV predates that of all other hantaviruses, including prototype HTNV, little is known about its biology and genetics. Whether TPMV is naturally harbored by the Asian house shrew or represents recent spillover from a rodent reservoir host is unknown. We present previously unpublished data on experimental TPMV infection in small laboratory animals. We also summarize information on the antigenic and phylogenetic relationships between TPMV and rodentborne hantaviruses that may cause hemorrhagic fever with renal syndrome (HFRS) (11) or hantavirus pulmonary syndrome (HPS) (12).
Shortly after TPMV was isolated, in vitro studies involving primary cultures of guinea pig embryonic kidney, lung, and heart cells supported replication of this virus (13). Although mild cytopathic effect was observed in these cell cultures, the kinetics of TPMV replication was not vigorously studied. Recent adaptation of the VRC-66412 strain of TPMV to the E6 clone of Vero cells (CRL 1586) showed no cytopathic effect. At a multiplicity of infection of 0.1, intracytoplasmic, virus-specific granular fluorescence appeared somewhat later in Vero E6 cells infected with TPMV than in those cells infected with HTNV or PUUV. Strains of HTNV isolated from striped field mice and strains of SEOV from Norway rats produce large plaques (6 mm diameter) on Vero-E6 cell monolayers stained 6 days after infection with neutral red. In contrast, the VRC-66412 strain of TPMV produces much smaller plaques (≈1–1.5 mm diameter) by staining with neutral red. These plaques are easily enumerated by immunohistochemical staining 12 days after infection.
In their primary rodent reservoir hosts, naturally occurring and experimentally induced hantavirus infections are subclinical and chronic (14–20). Experimental infection of striped field mice and bank voles with HTNV and PUUV, respectively, is characterized by transient viremia and short-lived shedding of virus in oropharyngeal secretions; prolonged excretion of virus in urine, feces, or both; and virus persistence in tissues, particularly lung (14,15,17–20). PUUV has been serially passaged only in laboratory-bred bank voles (17,19,20), and strains Hällnäs and K27 of PUUV cause an asymptomatic persistent infection in Mongolian gerbils (21) and Syrian hamsters (22), respectively. Horizontal intracage transmission has been demonstrated for HTNV and PUUV, but vertical transmission does not appear to occur (15,17,20,23). In infant mice and rats experimentally infected with HTNV and SEOV, respectively, fatal meningoencephalitis develops (24–27). In contrast, mice and rats >14–21 days of age are generally resistant to experimental HTNV and SEOV infection (26,27). Conversely, infant mice are resistant to experimental infection with PUUV (18,19), PHV (L.J. Baek, unpub. data), and SNV, the prototype sigmodontine rodentborne hantavirus that causes HPS (28).
Figure 1
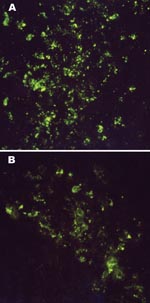
Figure 1. Intracytoplasmic virus-specific fluorescence in brain tissues of an 11-day-old Mongolian gerbil (A) and a 10-day-old NIH Swiss mouse (B) injected intracerebrally with 6,000 PFU of Thottapalayam virus (TPMV) strain VRC-66412 from...
To determine the host range of experimental TPMV infection and to ascertain whether susceptibility of small laboratory animals to disseminated TPMV infection is age-dependent, we infected NIH Swiss mice and Mongolian gerbils of different ages, as well as infant deer mice and gray short-tailed opossums (Monodelphis domestica), by the intracerebral route with 6,000 PFU of TPMV (Table 1). Infant Swiss NIH mice, deer mice, and gerbils were equally susceptible to fatal TPMV infection. Moreover, susceptibility to disseminated TPMV infection in NIH Swiss mice and gerbils was not age-dependent, as shown by lethal meningoencephalitis (characterized by hyperexcitability, ataxia, limb paralysis, and seizures) in animals infected at 1–21 days of age. TPMV antigen was detected in cryostat-cut sections of lung, brain, kidney, spleen, and liver of experimentally infected, moribund NIH Swiss mice and gerbils (Figure 1). Thus, unlike HTNV, SEOV, PUUV, PHV, and SNV, TPMV appears to have a much broader experimental host range in small laboratory animals.
Studies now in progress on experimental TPMV infection in laboratory-reared Asian house shrews should provide information about virus carriage and shedding. In addition, experimental demonstration of subclinical and chronic TPMV infection in Asian house shrews would also support the nonrodent reservoir host status.
The antigenic relationship of TPMV with 31 other hantavirus isolates has been investigated by ELISA and cross plaque-reduction neutralization tests (PRNTs) by using antisera from experimentally infected animals (9). Antisera prepared against strains of HTNV, SEOV, THAIV, PUUV, and PHV have 16-fold or lower ELISA titers to cell culture-derived TPMV antigen than to homologous antigen (9). ELISAs with monoclonal antibodies (MAbs) prepared against HTNV showed that certain epitopes defined by Gc-specific MAbs, but not Gn-specific MAbs, are conserved among most hantaviruses, including TPMV (9,29). Similarly, cross-immunoprecipitation of radionuclide-labeled TPMV and HTNV proteins have shown conserved N and Gc glycoprotein epitopes but not Gn epitopes (9). Of the 32 hantaviruses examined by PRNT, TPMV is the only virus that shows no cross-neutralization with any other hantavirus, i.e., none of the heterologous antisera neutralizes TPMV, and the antiserum to TPMV does not neutralize any other hantavirus.
Figure 2
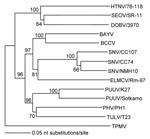
Figure 2. Phylogenetic relationship between Thottapalayam virus (TPMV) and other hantaviruses based on the nucleotide sequences of the full-length small (S) genomic segment, determined by using the neighbor-joining method. Numbers at each node...
Apart from being antigenically distinct, TPMV also appears to be the most genetically divergent member of the Hantavirus genus. Full-length S-segment nucleotide and deduced amino acid sequences have nearly the same calculated distances from all other hantaviruses, which suggests an early evolutionary divergence (Table 2). Phylogenetic analysis based on the N protein–encoding S segment, as determined by the maximum parsimony and neighbor-joining methods, supports this conclusion, in that TPMV is an outgroup and all other hantaviruses segregate into clades, which parallel the evolution of murid, arvicolid, and sigmodontine rodents (Figure 2). Further elucidation of the molecular phylogeny of TPMV has been hampered by the lack of TPMV M- and L-segment sequence information. After many failed attempts, the M- and L-genomic segments of TPMV have recently been fully sequenced (J.-W. Song, R. Yanagihara, unpub. data). Full-genome analysis of TPMV shows phylogenetic relationships with rodentborne hantaviruses, which are congruent with those formed only on the basis of the S segment.
Hantaviruses possess strikingly different degrees of pathogenicity for humans. Many viruses, particularly those harbored by arvicolid rodents, appear to be avirulent (such as PHV) or have low pathogenic potential (such as TULV). Among the HFRS- and HPS-causing pathogenic hantaviruses, differential use of β-3 integrins as cellular receptors on platelets and endothelial cells may account for vascular leakage and hemorrhage associated with HFRS and HPS (30,31). Preliminary studies indicate that TPMV, like PHV, uses β-1 rather than β-3 integrin (I.N. Gavrilovskaya, R. Yanagihara, unpub. data), which suggests that TPMV is nonpathogenic.
Immmunoglobulin G (IgG) against HTNV has been detected in sera from persons in southern India (32,33), but evidence for hantavirus disease in India is lacking. Nevertheless, because several species of Apodemus mice, including the wood mouse (A. sylvaticus), are present in India, the demonstrated seroreactivity to HTNV may represent cross-reactivity with another Apodemus-borne hantavirus. Alternatively, seroreactivity may indicate infection with a nonrodentborne hantavirus, such as TPMV. Because Asian house shrews are peridomestic, frequently living within or in close proximity to human dwellings, TPMV infection may occur in humans.
To begin to address this issue, researchers collected serum specimens from 363 life-long residents of Mumbai, India, during 1992 and 1993 as part of a study of retroviral infections, and tested them for serologic evidence of TPMV and SEOV infection by using the indirect immunofluorescent antibody technique. A total of 12 (3.3%) serum samples were reactive to TPMV (geometric mean titer 80.6), and 16 (4.4%) samples were reactive to SEOV (geometric mean titer 103.1). Attempts to verify the specificity of this immunoreactivity by PRNT were unsuccessful (J.-W. Song, unpub. data). More recently, however, evidence suggestive of TPMV infection was found in a Laotian immigrant with a febrile illness by using a Western immunoblot analysis and a newly developed ELISA. This ELISA used a recombinant TPMV N antigen that contained an E5/G6 epitope captured by MAb E5/G6 (29).
Although the detection of viruses in insectivores has been largely incidental or accidental, demonstration of Borna disease virus in brain tissues of the bicolored white-toothed shrew (Crocidura leucodon) (34) suggests that insectivores may play a greater role in the ecology of zoonotic diseases than previously appreciated. The prototype shrewborne hantavirus, TPMV, must be viewed within this context. Although limited data do not indicate that TPMV is a human pathogen, other shrewborne hantaviruses may be pathogenic for humans. In this regard, no one had the prescience to predict that hantaviruses harbored by sigmodontine rodents would be etiologically associated with an acute, rapidly progressive, frequently fatal respiratory illness in the Americas, now known as HPS. The realization that rodentborne hantaviruses are capable of causing diseases as clinically disparate as HFRS and HPS increases the possibility that hantaviruses harbored by nonrodent hosts may similarly cause a wide spectrum of febrile diseases or be linked with a syndrome currently of unknown etiology. Development of reagents directed toward insectivore serum proteins would greatly increase the sensitivity and specificity of serologic assays to ascertain antihantaviral immunologic responses in shrews and result in improved screening for new shrewborne hantaviruses.
Even in the absence of such reagents, several insectivore species are already known to be prime candidates for intensive investigations aimed at identifying new hantaviruses and exploring their disease associations. As examples, hantavirus antigens have been previously detected in tissues of the Eurasian common shrew (Sorex araneus), Eurasian water shrew (Neomys fodiens), and common mole (Talpa europea) in the former Soviet Union (35,36), and seroreactivity suggestive of hantavirus infection was found in short-tailed shrew (Blarina brevicauda) in the United States (37). In addition, hantaviruses isolated more than 2 decades ago from the greater white-toothed shrew (Crocidura russula) (38) and Chinese mole shrew (Anourosorex squamipes) (38) in Sichuan Province, People’s Republic of China, have been inadequately characterized. The probability is high that >1 of these shrewborne hantaviruses may be phylogenetically distinct.
Figure 3
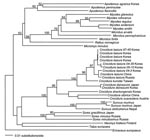
Figure 3. Phylogenetic relationship between Suncus murinus and other insectivores and rodents in a 401-nt cytochrome b region of mitochondrial DNA determined by using the neighbor-joining method. Numbers at each node are bootstrap...
Another approach to the targeted discovery of new hantaviruses harbored by shrews relies on molecular phylogeny. By constructing phylogenetic trees based on mitochondrial or nuclear gene DNA sequences, existence of hantaviruses in the Korean field mouse (A. peninsulae) (39) and the royal vole (Myodes regulus, formerly Eothenomys regulus) (J.-W. Song, unpub. data) was correctly predicted. When this predictive paradigm is applied to insectivores, species of Crocidura and Sorex genera would be expected to serve as reservoir hosts of hantaviruses because of their close phylogenetic proximity to S. murinus (Figure 3). The recent detection of a novel hantavirus in the Therese shrew (Crocidura theresae) in Guinea (40) supports this conjecture. In addition, aided by primers based on the complete genome of TPMV, new hantaviruses have been found in 4 shrew species in the family Soricidae from Eurasia and the Americas (J.-W. Song, R. Yanagihara, unpub. data, and S. Arai, R. Yanagihara, unpub. data). These newly identified shrewborne hantaviruses provide new knowledge about the genetic diversity of hantaviruses as well as possible insights into their evolutionary origin through host-switching events.
Fundamental to the discovery and characterization of new hantaviruses, whether harbored by insectivores or rodents, is their relevance to human health. Because insectivore populations are generally much smaller than rodent populations, the probability of contact between humans and most insectivore species (and their excretions) may be too low for virus transmission. However, this probability is true for most zoonotic microbes, which only rarely infect humans. Thus, in the absence of disease outbreaks, zoonotic diseases frequently go unrecognized. In this regard, HPS would have similarly gone undetected had cases not clustered in time and space and had a closely knit group of dedicated and astute healthcare workers not recognized that something unusual was happening. The long-awaited clue of finding IgG against TPMV in a febrile Laotian immigrant (29) might indicate cross-reactivity to a pathogenic shrewborne hantavirus in Southeast Asia.
Dr Song is professor of microbiology at Korea University. His research interests include global epizootiology and epidemiology of hantaviruses.
Acknowledgments
We thank Allison Toney for her contributions to this study while she held a National Research Council Research Associateship at the US Army Medical Research Institute of Infectious Diseases.
This study was supported in part by grants from the Ministry of Science and Technology (Korea Science and Engineering Foundation), Korea (no. R21-2005-000-10017-0) and the National Center for Research Resources, National Institutes of Health (P20RR018727).
References
- Schmaljohn CS, Dalrymple JM. Analysis of Hantaan virus RNA: evidence for a new genus of Bunyaviridae. Virology. 1983;131:482–91. DOIPubMedGoogle Scholar
- Schmaljohn CS, Hasty SE, Harrison SA, Dalrymple JM. Characterization of Hantaan virions, the prototype virus of hemorrhagic fever with renal syndrome. J Infect Dis. 1983;148:1005–12.PubMedGoogle Scholar
- Schmaljohn CS, Hasty SE, Dalrymple JM, LeDuc JW, Lee HW, von Bonsdorff CH, Antigenic and genetic properties of viruses linked to hemorrhagic fever with renal syndrome. Science. 1985;227:1041–4. DOIPubMedGoogle Scholar
- Plyusnin A, Vapalahti O, Vaheri A. Hantaviruses: genome structure, expression and evolution. J Gen Virol. 1996;77:2677–87. DOIPubMedGoogle Scholar
- Vapalahti O, Mustonen J, Lundkvist A, Henttonen H, Plyusnin A, Vaheri A. Hantavirus infections in Europe. Lancet Infect Dis. 2003;3:653–61. DOIPubMedGoogle Scholar
- Klempa B, Fichet-Calvet E, Lecompte E, Auste B, Aniskin V, Meisel H, Hantavirus in African wood mouse, Guinea. Emerg Infect Dis. 2006;12:838–40.PubMedGoogle Scholar
- Carey DE, Reuben R, Panicker KN, Shope RE, Myers RM. Thottapalayam virus: a presumptive arbovirus isolated from a shrew in India. Indian J Med Res. 1971;59:1758–60.PubMedGoogle Scholar
- Zeller HG, Karabatsos N, Calisher CH, Digoutte J-P, Cropp CB, Murphy FA, Electron microscopic and antigenic studies of uncharacterized viruses. II. Evidence suggesting the placement of viruses in the family Bunyaviridae. Arch Virol. 1989;108:211–27. DOIPubMedGoogle Scholar
- Chu YK, Lee HW, LeDuc JW, Schmaljohn CS, Dalrymple JM. Serological relationships among viruses in the Hantavirus genus, family Bunyaviridae. Virology. 1994;198:196–204. DOIPubMedGoogle Scholar
- Xiao S-Y, LeDuc JW, Chu YK, Schmaljohn CS. Phylogenetic analyses of virus isolates in the genus Hantavirus, family Bunyaviridae. Virology. 1994;198:205–17. DOIPubMedGoogle Scholar
- Yanagihara R, Gajdusek DC. Hemorrhagic fever with renal syndrome: a historical perspective and review of recent advances. In: Gear JHS, editor. CRC handbook of viral and rickettsial hemorrhagic fevers. Boca Raton (FL): CRC Press; 1988. p. 151–88.
- Nichol ST, Spiropoulou CF, Morzunov S, Rollin PE, Ksiazek TG, Feldmann H, Genetic identification of a hantavirus associated with an outbreak of acute respiratory illness. Science. 1993;262:914–7. DOIPubMedGoogle Scholar
- Venkateshan CN. Studies on the susceptibility of guinea pig embryonic cell cultures to some arboviruses of India. In: Abstracts of the Fifteenth Annual Conference of Microbiologists of India. Bangalore: Indian Institute of Science; 1974.
- Lee HW, French GR, Lee P-W, Baek LJ, Tsuchiya K, Foulke RS. Observations on natural and laboratory infection of rodents with the etiologic agent of Korean hemorrhagic fever. Am J Trop Med Hyg. 1981;30:477–82.PubMedGoogle Scholar
- Lee HW, Lee P-W, Baek LJ, Song CK, Seong IW. Intraspecific transmission of Hantaan virus, the etiologic agent of Korean hemorrhagic fever, in the rodent Apodemus agrarius. Am J Trop Med Hyg. 1981;30:1106–12.PubMedGoogle Scholar
- Lee HW, Baek LJ, Johnson KM. Isolation of Hantaan virus, the etiologic agent of Korean hemorrhagic fever, from wild urban rats. J Infect Dis. 1982;146:638–44.PubMedGoogle Scholar
- Brummer-Korvenkontio M, Henttonen H, Vaheri A. Hemorrhagic fever with renal syndrome in Finland: ecology and virology of nephropathia epidemica. Scand J Infect Dis Suppl. 1982;36:88–91.PubMedGoogle Scholar
- Gavrilovskaya IN, Chumakov MP, Apekina NS, Ryltseva EV, Martiyanova LI, Gorbachkova EA, Adaptation to laboratory and wild animals of the haemorrhagic fever with renal syndrome virus present in the foci of European U.S.S.R. Brief report. Arch Virol. 1983;77:87–90. DOIPubMedGoogle Scholar
- Yanagihara R, Svedmyr A, Amyx HL, Lee P-W, Goldgaber D, Gajdusek DC, Isolation and propagation of nephropathia epidemica virus in bank voles. Scand J Infect Dis. 1984;16:225–8. DOIPubMedGoogle Scholar
- Yanagihara R, Amyx HL, Gajdusek DC. Experimental infection with Puumala virus, the etiologic agent of nephropathia epidemica, in bank voles (Clethrionomys glareolus). J Virol. 1985;55:34–8.PubMedGoogle Scholar
- Yanagihara R, Goldgaber D, Gajdusek DC. Propagation of nephropathia epidemica virus in Mongolian gerbils. J Virol. 1985;53:973–5.PubMedGoogle Scholar
- Chu YK, Jennings GB, Schmaljohn CS. A vaccinia virus-vectored Hantaan virus vaccine protects hamsters from challenge with Hantaan and Seoul viruses but not Puumala virus. J Virol. 1995;69:6417–23.PubMedGoogle Scholar
- Lee P-W, Yanagihara R, Gibbs CJ Jr, Gajdusek DC. Pathogenesis of experimental Hantaan virus infection in laboratory rats. Arch Virol. 1986;88:57–66. DOIPubMedGoogle Scholar
- Tsai TF, Bauer S, McCormick JB, Kurata T. Intracerebral inoculation of suckling mice with Hantaan virus. Lancet. 1982;2:503–4. DOIPubMedGoogle Scholar
- Kurata T, Tsai TF, Bauer SP, McCormick JB. Immunofluorescence studies of disseminated Hantaan virus infection of suckling mice. Infect Immun. 1983;41:391–8.PubMedGoogle Scholar
- Yamanouchi T, Domae K, Tanishita O, Takahashi Y, Yamanishi K, Takahashi M, Experimental infection in newborn mice and rats by hemorrhagic fever with renal syndrome (HFRS) virus. Microbiol Immunol. 1984;28:1345–53.PubMedGoogle Scholar
- Nakamura T, Yanagihara R, Gibbs CJ Jr, Amyx HL, Gajdusek DC. Differential susceptibility and resistance of immunocompetent and immunodeficient mice to fatal Hantaan virus infection. Arch Virol. 1985;86:109–20. DOIPubMedGoogle Scholar
- Elliott LH, Ksiazek TG, Rollin PE, Spiropoulou CF, Morzunov S, Monroe M, Isolation of the causative agent of hantavirus pulmonary syndrome. Am J Trop Med Hyg. 1994;51:102–8.PubMedGoogle Scholar
- Okumura M, Yoshimatsu K, Kumperasart S, Nakamura I, Ogino M, Taruishi M, Development of serological assays for Thottapalayam virus, an insectivore-borne hantavirus. Clin Vaccine Immunol. 2007;14:173–81. DOIPubMedGoogle Scholar
- Gavrilovskaya IN, Shepley M, Shaw R, Ginsberg MH, Mackow ER. beta3 integrins mediate the cellular entry of hantaviruses that cause respiratory failure. Proc Natl Acad Sci U S A. 1998;95:7074–9. DOIPubMedGoogle Scholar
- Gavrilovskaya IN, Brown EJ, Ginsberg MH, Mackow ER. Cellular entry of hantaviruses which cause hemorrhagic fever with renal syndrome is mediated by beta3 integrins. J Virol. 1999;73:3951–9.PubMedGoogle Scholar
- Lee PW, Gibbs CJ Jr, Gajdusek DC, Svedmyr A. Antibody to Korean haemorrhagic fever virus in man in parts of the world where haemorrhagic fever with renal syndrome is not known. Lancet. 1981;2:256–7.PubMedGoogle Scholar
- Chandy S, Mitra S, Sathish N, Vijayakumar TS, Abraham OC, Jesudason MV, A pilot study for serological evidence of hantavirus infection in human population in south India. Indian J Med Res. 2005;122:211–5.PubMedGoogle Scholar
- Hilbe M, Herrsche R, Kolodziejek J, Nowotny N, Zlinszky K, Ehrensperger F. Shrews as reservoir hosts of Borna disease virus. Emerg Infect Dis. 2006;12:675–7.PubMedGoogle Scholar
- Gavrilovskaya IN, Apekina NS, Myasnikov YA, Bernshtein AD, Ryltseva EV, Gorbachkova EA, Features of circulation of hemorrhagic fever with renal syndrome (HFRS) virus among small mammals in the European U.S.S.R. Arch Virol. 1983;75:313–6. DOIPubMedGoogle Scholar
- Tkachenko EA, Ivanov AP, Donets MA, Miasnikov YA, Ryltseva EV, Gaponova LK, Potential reservoir and vectors of haemorrhagic fever with renal syndrome (HFRS) in the U.S.S.R. Ann Soc Belg Med Trop. 1983;63:267–9.PubMedGoogle Scholar
- Lee P-W, Amyx HL, Yanagihara R, Gajdusek DC, Goldgaber D, Gibbs CJ Jr. Partial characterization of Prospect Hill virus isolated from meadow voles in the United States. J Infect Dis. 1985;152:826–9.PubMedGoogle Scholar
- Yan DY, Xie YJ, Zhang CA, McCormick JB, Sanchez A, Engelman HM, New isolates of HFRS virus in Sichuan, China, and characterization of antigenic differences by monoclonal antibodies. Lancet. 1986;1:1328. DOIPubMedGoogle Scholar
- Baek LJ, Kariwa H, Lokugamage K, Yoshimatsu K, Arikawa J, Takashima I, Soochong virus: a genetically distinct hantavirus isolated from Apodemus peninsulae in Korea. J Med Virol. 2006;78:290–7. DOIPubMedGoogle Scholar
- Klempa B, Fichet-Calvet E, Lecompte E, Auste B, Aniskin V, Meisel H, Novel hantavirus sequences in shrew, Guinea. Emerg Infect Dis. 2007;13:520–2. [cited 2007 Apr 23]. Available from http://www.cdc.gov/eid/content/13/3/520B.htm
Figures
Tables
Cite This ArticleTable of Contents – Volume 13, Number 7—July 2007
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Richard Yanagihara, Pacific Center for Emerging Infectious Diseases Research, John A. Burns School of Medicine, University of Hawaii at Manoa, 651 Ilalo St, BSB 320L, Honolulu, HI 96813, USA;
Top