Volume 13, Number 7—July 2007
Research
Virulence Characteristics of Klebsiella and Clinical Manifestations of K. pneumoniae Bloodstream Infections
Cite This Article
Citation for Media
Abstract
We studied 455 consecutive episodes of Klebsiella pneumoniae bacteremia occurring in 7 countries. Community-acquired pneumonia and an invasive syndrome of liver abscess, meningitis, or endophthalmitis occurred only in Taiwan and South Africa. Infections by K1 and K2 capsular serotype, the mucoid phenotype, and aerobactin production were important determinants of virulence. The mucoid phenotype was seen in 94% of isolates in patients with community-acquired pneumonia and in 100% of isolates that caused the invasive syndrome in Taiwan and South Africa, compared with only 2% of isolates elsewhere. Mortality of mice injected with mucoid strains (69%) was strikingly higher than that occurring in mice injected with nonmucoid strains (3%, p<0.001). Differences in clinical features of bacteremic infection with K. pneumoniae are due to the virulence factors expressed by the organism.
In the past decade, geographic differences have been recognized in the spectrum of disease caused by Klebsiella pneumoniae. These differences include a preponderance of severe invasive disease in Taiwan and other parts of Asia (1–8). A characteristic syndrome has emerged in which liver abscess is accompanied by K. pneumoniae bacteremia and sometimes by endophthalmitis or meningitis. This is typically a community-acquired infection that occurs in patients with diabetes mellitus. Reports of this syndrome from North America, Europe, and Australia are uncommon (2).
Additionally, K. pneumoniae has long been recognized as a possible cause of community-acquired pneumonia. Over the past 2 decades, K. pneumoniae has been an exceedingly rare cause of community-acquired pneumonia in North America, Europe, and Australia (2,9,10). Yet, it remains an important cause of severe community-acquired pneumonia in Asia and Africa (11–15). In these regions, patients also have classic risk factor of alcoholism (2).
We have completed a prospective study of 455 patients from 7 countries with K. pneumoniae bacteremia (2). We found that although nosocomial infections with K. pneumoniae occurred worldwide, some manifestations of community-acquired infection (namely, liver abscess and community-acquired pneumonia) were geographically restricted. These manifestations of disease occurred almost exclusively in Taiwan and South Africa (2). Potential explanations for these geographic differences in clinical manifestations include host factors such as rates of diabetes mellitus, alcoholism, access to healthcare, and socioeconomic factors.
Another explanation for these differences is related to the organism. In this study, we performed capsular serotyping, determined the presence of mucoid phenotype and aerobactin production, and assessed lethality in a murine model and correlated these in vitro and in vivo results with the clinical manifestation of patients with K. pneumoniae bloodstream infections. Our aim was to determine whether the different manifestations of infection occurring in different geographic regions could be correlated with differences in organism characteristics.
Study Design
A prospective, observational study of consecutive, sequentially encountered patients with K. pneumoniae bacteremia was conducted in 12 hospitals in the United States, Taiwan, Australia, South Africa, Turkey, Belgium, and Argentina. No patients were excluded from analysis. The study period was January 1, 1996, to December 31, 1997. Patients >16 years of age with positive blood cultures for K. pneumoniae were enrolled and completed a 188-item study form. Patients were followed up for 1 month after the onset of bacteremia to assess clinical outcome, including deaths and infectious complications. The study was observational in that administration of antimicrobial agents and other therapeutic management were controlled by the patient’s physician, not the investigators. The study was approved by institutional review boards as required by local hospital policy.
Definitions
Terms were defined a priori (that is, before data analysis). Community-acquired bacteremia was defined as a positive blood culture taken on admission or within 48 hours of admission. Site of infection accompanying the bacteremia was determined as pneumonia, urinary tract infection, meningitis, incisional wound infection, other soft tissue infection, intraabdominal infection, and primary bloodstream infection by using Centers for Disease Control and Prevention definitions (16). “Invasive” infections accompanying K. pneumoniae bacteremia were further defined as liver abscess, meningitis, or endophthalmitis. Liver abscess was defined by the coexistence of blood cultures positive for K. pneumoniae and evidence of an intrahepatic abscess cavity by ultrasonography or computed tomography. Meningitis was defined as culture of K. pneumoniae from the cerebrospinal fluid. Endophthalmitis was defined as decreased visual acuity, pain, hypopyon, or severe anterior uveitis concurrent with K. pneumoniae bacteremia in a patient.
Microbiology
Blood culture isolates of K. pneumoniae were sent by the participating hospitals on nutrient agar slants to the Special Pathogens Laboratory in Pittsburgh. There, the identity of each isolate as K. pneumoniae was confirmed by using the Vitek GNI system (bioMérieux Vitek, Hazelwood, MO, USA).
Figure 1

Figure 1. Mucoid phenotype of Klebsiella pneumoniae. When colonies were touched with a loop and the loop lifted vertically from the surface of the agar plate, mucoid isolates adhered to the loop as...
The isolates were classified phenotypically as mucoid or nonmucoid. Colonies were touched with a loop; the loop was then lifted vertically from the surface of the agar plate. Mucoid phenotype was defined as being present when a stringlike growth was observed to attach to the loop as it was lifted from the plate (Figure 1). Presence of the rmpA gene (rmp = regulator of the mucoid phenotype) was sought by DNA dot blot hybridization by using a 640-bp probe (position 478–1117 of the rmpA gene; accession no. X17518). The probe was produced by direct digoxigenin (DIG)-labeled PCR by using the PCR DIG probe synthesis kit (Roche Diagnostics, Basel, Switzerland) and the primers Kleb_MP_F1 (5′-GAG CAA AGT TAC TGT TTC TAT GGA-3′) and Kleb_MP-R1 (5′-TGA GCC ATC TTT CAT CAA CC-3′) on the K. pneumoniae strain B 5055. Dot blot hybridization was performed according to the manufacturer’s protocol on Hybond N+ nylon membranes (Amersham, Pharmacia Biotech, Piscataway, NJ, USA), and the hybridized probe was visualized by using the DIG nucleic acid detection kit (Roche Diagnostics).
Capsular K serotyping was performed at the World Health Organization International Escherichia and Klebsiella Reference Centre (Copenhagen, Denmark) by using standard methods. In brief, K-typing was conducted by counter current immunoelectrophoresis (CCIE) with a modified version of the method described by Palfreyman (17). An extract was used as antigen instead of a whole cell suspension; the extract use was a modification because it was only heated once for 1 h at 100°C before centrifugation (18). All isolates with negative or doubtful reactions in CCIE were investigated by the classic Quellung technique, and K-type nontypeable isolates were investigated for the presence (K+) or absence (K–) of a visible capsule by wet mount microscopy with India ink. Lipopolysaccharide O typing was performed by a previously described inhibition ELISA (19).
Aerobactin production was demonstrated by a cross-feeding bioassay that used Escherichia coli strain LG 1522 (20). The clinical isolates were grown overnight in M9 broth containing the iron chelator 2-2′ dipyridyl. Strains were spotted onto hardened dipyridyl minimal agar plates. After 18 hours’ incubation at 37°C, satellite growth of the indicator strain LG 1522 around the spots indicated aerobactin production.
Lethality in Mice
A standard inoculum of 1–2 × 107 bacteria in the logarithmic phase of growth from blood culture isolates from each study site was injected intravenously into the tail vein of C57/BL6J black, female mice, 8–12 weeks old. Two mice (Jackson Laboratories, Bar Harbor, ME, USA) were inoculated with each strain. Mortality of the mice was observed at 24 hours postinjection. The animal experiments were approved by the Institutional Review Board of the Veterans Affairs Medical Center, Pittsburgh, Pennsylvania, USA.
Pulsed-Field Gel Electrophoresis (PFGE)
The genotypic relationships of K. pneumoniae bloodstream isolates were determined by using PFGE. PFGE was performed by means of the CHEF-DR II system (Bio-Rad, Richmond, CA, USA) with use of the restriction endonuclease Xba I (New England Biolabs, Beverly, MA, USA). DNA was subjected to electrophoresis for 22 hours at 14°C in a 1% agarose gel at 6 V/cm with a linear gradient pulse time of 5–35 seconds. The gels were analyzed by using the Gel Doc 2000 software (Bio-Rad).
Statistics
Patient demographics and laboratory data were entered into PROPHET Statistics version 6.0 (AbTech Corporation, Charlottesville, VA, USA). The χ2 or Fisher test was used to compare categorical variables. Continuous variables were compared by using the t test or the Mann-Whitney test. Multivariate analysis was used to determine which risk factors for mouse lethality by univariate analysis were independently significant.
Serotypes of K. pneumoniae Bacteremic Strains
During the study period, 455 episodes of K. pneumoniae bloodstream infection occurred; 141 community-acquired bloodstream isolates were available and were tested for K serotype. Three isolates were not encapsulated, and 20 were nontypeable. Forty-seven different capsular serotypes were found in the remaining strains; K1 (16%; 23/141), K2 (11%; 16/141), and K54 (9%; 12/141) were the most common serotypes.
One hundred percent (23/23) of K1 serotype strains, 94% (15/16) of K2 serotype strains, and 100% (12/12) of K54 serotype strains were from Taiwan or South Africa. Forty-seven percent (23/49) of isolates from patients with community-acquired pneumonia and 50% (7/14) of isolates from patients with invasive syndromes possessed the K1 or K2 serotype. In comparison, 12% (9/78) isolates from patients with other manifestations of community-acquired K. pneumoniae bacteremia had the K1 or K2 serotype (p<0.001).
Seventy-three randomly chosen hospital-acquired bloodstream isolates from the same multinational study were tested for K serotype. One isolate was not encapsulated, and 5 isolates were nontypeable. In contrast to the predominance of K1 serotypes in community-acquired infections, 1% (1/75) of hospital-acquired isolates were K1 (p = 0.0015). The percentage of K2 serotypes in hospital acquired isolates (9%, 7/75) was similar to that observed in community-acquired isolates (11%, 16/141) (p = 0.76). The hospital-acquired K1 isolate was from Taiwan; 4 hospital-acquired K2 isolates were from Africa, 2 were from the United States, and 1 was from Taiwan. In contrast to the predominance of K1 and K2 serotypes in community-acquired pneumonia (47%, 23/49 isolates), these serotypes were significantly less likely to occur in nosocomial pneumonia (11%, 2/18 cases; p = 0.022).
Relationship between Capsular Serotype and Lethality in Mice
When community-acquired strains only were examined, highest lethality was observed with strains of K1 serotype (mouse deaths 65%) and K2 serotype (mouse deaths 81%). In contrast, lethality for community-acquired strains of other serotypes was significantly lower (23% died; p<0.001). The proportion of deaths in mice injected with community-acquired strains of serotype K54 was 9%.
When hospital-acquired strains were examined, mouse mortality was only 8%. The only lethal strains were K1 and K17 serotypes. None of the 7 hospital-acquired strains of serotype K2 proved lethal to mice.
When both community- and hospital-acquired strains were assessed together, lethality was significantly higher for strains of K1 (mouse deaths 67%) and K2 serotype (mouse deaths 59%) than for strains of other serotype (mouse deaths 20%) (p<0.001). The lethality of all strains of serotype 54 was 12.5%.
Lipopolysaccharide O Typing
Of 195 isolates which underwent O typing, 30% (58/195) were group O1, 17% (34/195) were O2, 8% (16/195) were O3, 2% (3/195) were O4, 3% (6/195) were O5, 1% (2/195) were O2ac, 7% (13/195) were O–, and 32% (63/195) were O+ but not typable. No strains of O group O3, O4, O5, O2ac or O– were lethal to mice. In contrast, 21% (12/58) of O1 strains, 26% (9/34) of O2 strains, and 37% (23/63) of nontypeable O+ strains were lethal to mice.
Relationship between Mucoid Phenotype and Type of Infection
The mucoid phenotype was observed in 93% (13/14) of strains from patients with invasive disease, in 67% (33/49) of strains from patients with community-acquired pneumonia, and in 28% (22/78) of strains from patients with other sites of community-acquired infection (p<0.001). Mucoid strains of community-acquired bacteremic K. pneumoniae were exclusively found in Taiwan and South Africa; 56% (45/80) and 60% (21/35) of Taiwanese and African strains, respectively, were mucoid, whereas no mucoid strains were detected in community-acquired bacteremic strains from other countries (Table 1). In Taiwan and South Africa, mucoid strains predominated in community-acquired strains (57%, 66/115) compared with hospital-acquired strains (18%, 7/38) (Table 2).
In Taiwan and South Africa, community-acquired pneumonia was due to mucoid strains of K. pneumoniae in younger patients with no serious underlying disease, while nonmucoid strains predominated in older patients and those with serious underlying disease (Table 3). In both countries combined, 94% (29/31) strains from community-acquired pneumonia patients with no serious underlying disease had the mucoid phenotype compared to 19% (3/16) strains from patients with serious underlying disease (p<0.001). All isolates (13/13) from patients in Taiwan or South Africa with the invasive syndrome of liver abscess, meningitis, or endophthalmitis had the mucoid phenotype (Table 4).
The proportion of deaths in mice injected with mucoid strains (69% of mice died) was strikingly higher than that occurring in mice injected with nonmucoid strains (3% mice died) (p<0.001). There was no association between human deaths and the presence of a mucoid strain (39% died) or a nonmucoid strain (35% died) (p>0.20). In a multivariate model, increased severity of illness score when first evaluated (p = 0.0001), but not infection with a mucoid strain, country of origin, or history of alcoholism (p>0.20 for all) was associated with human deaths.
Association between Phenotypic Evidence of Mucoidity and Presence of rmpA Gene
Phenotypic evidence of mucoidity as judged by the definition in the methods section (“a string-like growth observed to attach to the loop as it was lifted from the plate”) (Figure 1) was highly correlated with the presence of the rmpA gene. Of 77 mucoid isolates, 86% (66/77) were rmpA gene positive, and 14% (11/77) were rmpA gene negative. Of 137 nonmucoid isolates, 93% (128/137) were negative for the rmpA gene and 7% (9/137) were rmpA positive.
Relationship between Aerobactin Production and Type of Infection
The presence of the rmpA gene and phenotypic evidence of aerobactin production were closely correlated. Ninety-six percent of rmpA gene–positive isolates were aerobactin producers; aerobactin was produced by 2% of isolates that were rmpA gene–negative.
Associations between aerobactin production and type of infection were similar to those between the mucoid phenotype and type of infection. Only 6% (4/62) strains from patients in countries other than Taiwan and South Africa were aerobactin producers. In Taiwan and South Africa, 66% of patients with community-acquired pneumonia and 85% of patients with the invasive syndrome had aerobactin-producing strains, in comparison with 42% of patients with other community-acquired infections and 16% of patients with hospital-acquired strains (Table 2).
Relationship between Mucoid Phenotype, Capsular Serotype, and Lethality in Mice
When both community-acquired and hospital-acquired strains were considered together, 77% (36/47) of isolates of serotypes K1 and K2 were found to be mucoid. Of the other 45 serotypes, 24% (40/167) were mucoid (p<0.001).
However, none of the mice inoculated with nonmucoid K1 or K2 serotype strains died, compared with 81% of mice inoculated with mucoid K1 or K2 serotype strains (p<0.001). Just 4% of mice inoculated with nonmucoid strains of serotypes other than K1 or K2 died, compared with 44% mice inoculated with mucoid organisms of serotypes other than K1 or K2 (p<0.001).
When the parameters of mucoid phenotype, serotypes K1 and K2, and country of origin were assessed in the multivariate model of lethality to mice, mucoid phenotype was strongly associated with the death of mice (p = 0.001). Presence of serotypes K1 and K2 approached statistical significance (p = 0.05).
PFGE
Figure 2

Figure 2. Pulsed-field gel electrophoresis of bacteremic Klebsiella pneumoniae isolates of serotype K1.
Figure 3
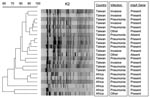
Figure 3. Pulsed-field gel electrophoresis of bacteremic Klebsiella pneumoniae isolates of serotype K2.
PFGE was performed on strains of the same serotype. Dendrograms of organisms of serotype K1 are shown in Figure 2 and dendrograms of serotype K2 in Figure 3.
We have been able to evaluate geographic differences in community-acquired K. pneumoniae infections by studying consecutive patients with community-acquired K. pneumoniae bacteremia from 7 different countries during the same period. It could be hypothesized that patient characteristics are primarily responsible for these differences. For example, genetic predilections (susceptibility of Asians to liver abscess), underlying diseases (for example, higher prevalence of chronic hepatitis B virus infection in Taiwan), social factors (different foods or cultural practices), and economic factors (for example, access to healthcare, antimicrobial drug usage) may be responsible for the different manifestations of serious K. pneumoniae infection observed in different regions. Despite these other possibilities, our experimental studies suggest that the differences in clinical features arise from differences in the virulence of individual microorganisms.
In particular, we found that strains with K1 or K2 serotype, strains with a mucoid phenotype, and strains that are capable of aerobactin production are rarely found to cause substantial infection in patients from study hospitals outside Taiwan and South Africa. Strains with such virulence characteristics were more likely to cause community-acquired infections than hospital-acquired infections. When strains with these virulence characteristics were inoculated into mice, deaths exceeded 80% compared with mortality rates of <5% in mice inoculated with strains lacking these characteristics.
Additionally, by PFGE, we found genetically related strains possessing all 3 virulence characteristics (Figures 2, 3). Taiwanese investigators have debated whether K. pneumoniae strains that cause liver abscess in Taiwan are clonally related (1,4,6,7,21). Genetic relatedness in K. pneumoniae strains that cause community-acquired pneumonia has not been previously described. However, we have found that genotypically related organisms were responsible for bacteremic community-acquired pneumonia due to K. pneumoniae and sometimes both pneumonia and liver abscess or meningitis. Whether K. pneumoniae is spread from person to person, whether related strains are acquired from common sources, or whether virulent strains arise from a common ancestor remains to be determined.
Much prominence has been placed in the past on the role of capsule in the pathogenesis of K. pneumoniae infections. Capsular types K1 and K2 have been regarded as particularly virulent (22). Taiwanese researchers have found that serotype K1 is frequently associated with community-acquired K. pneumoniae bacteremia (3,23). We have also confirmed that serotypes K1 and K2 occur more frequently in isolates from community-acquired infections in Taiwan and South Africa than from hospital-acquired isolates in these countries or elsewhere (Table 1).
The degree of virulence conferred by a particular K antigen may be related to the mannose content of the capsular polysaccharide. Capsular types with high virulence in animal models (for example, K2) lack mannose-α-2/3-mannose structures found in capsular types of lower virulence (24). The mannose-α-2/3-mannose structures are recognized by a surface lectin of macrophages, which mediate complement and antibody-independent phagocytosis. Strains that lack these sequences (for example, those with the K2 antigen) may not be recognized by macrophages, and hence phagocytosis may not take place. Furthermore, surfactant protein A (the main protein component of lung surfactant) enhances the phagocytosis by alveolar macrophages of strains that bear mannose-α-2/3-mannose structures in their capsule, but not strains which lack the mannose structure (25).
Some strains belonging to the K2 serotype are not as virulent as others (26). Thus, factors other than capsule may also be important for virulence. Although previous authors did not find any markers for these differences (26), we found that mucoid strains of K1 or K2 serotype were more virulent to mice than nonmucoid strains of the same serotype. No mouse inoculated with nonmucoid K1 or K2 serotype strains died, compared to 81% mice inoculated with mucoid K1 or K2 serotype strains (p<0.001). Multivariate analysis of variables related to mouse mortality rates showed that mucoidity was more closely associated with death than was capsular serotype. Contrary to popular belief, the biochemical nature of the mucoid phenotype may be unrelated to capsular polysaccharide but rather related to extracapsular polysaccharide (27). A previous study has shown that the mucoid phenotype may be due to a gene designated rmpA (regulator of mucoid phenotype) (28). In another mouse model, a mutant carrying this gene was 1,000-fold more virulent than an isolate without the gene. Extracellular polysaccharides may protect mucoid strains of K. pneumoniae from phagocytosis by neutrophils and from serum killing by complements (27).
We found that the mucoid phenotype frequently coexists with aerobactin production. The growth of bacteria in host tissues is limited not only by host defense mechanisms but also by its supply of available iron. The supply of free iron in the host milieu may be extremely low; many bacteria attempt to secure their supply of iron in the host by secreting high-affinity iron chelators called siderophores. Aerobactin is a hydroxamate-type siderophore occasionally found in Klebsiella strains. K. pneumoniae strains that produce aerobactin were more virulent in our mouse model, whereas strains not producing this siderophore were less likely to be; additionally, patients with severe community-acquired infection were more likely to be infected by aerobactin-producing strains. In another mouse model, transfer of a recombinant plasmid harboring the genes for aerobactin and its receptor enhanced the virulence of an otherwise avirulent strain by 100-fold (29). The strong association found in this study between mucoid phenotype (rmpA positive isolates) and aerobactin production suggests that the 2 virulence characteristics might be genetically coupled on a large virulence plasmid, as has previously been demonstrated (28). We have not yet determined which of these 2 virulence factors is more important.
Strains harboring these virulence factors appear to be more frequent in certain geographic regions, and this may explain geographic differences in manifestation of community-acquired Klebsiella infections. The evolutionary genetics of K. pneumoniae have never been explored. To our knowledge, we have been the first to find clones bearing multiple virulence characteristics that are responsible for life-threatening community-acquired K. pneumoniae pneumonia in otherwise healthy persons. Investigation into the mechanisms of virulence in K. pneumoniae could lead to preventive measures (such as vaccination) in high-risk parts of the world.
Dr Yu is professor of medicine at the University of Pittsburgh. His research interests include Legionnaires’ disease, pneumonia, and antibiotic-resistant bacteria.
Acknowledgments
This article is dedicated to the memory of Vicente J. Benedi, who died unexpectedly in December 2002. We thank Feng-Yee Chang for his expert review and suggested additions to the manuscript.
Merck and Company provided partial support for the laboratory studies but played no role in study design, interpretation, or approval of the study. Keith P. Klugman is a consultant for Abbott Laboratories, Astra-Zeneca, Bayer, and GlaxoSmithKline.
References
- Cheng HP, Chang FY, Fung CP, Siu LK. Klebsiella pneumoniae liver abscess in Taiwan is not caused by a clonal spread strain. J Microbiol Immunol Infect. 2002;35:85–8.PubMedGoogle Scholar
- Ko WC, Paterson DL, Sagnimeni AJ, Hansen DS, Von Gottberg A, Mohapatra S, Community-acquired Klebsiella pneumoniae bacteremia: global differences in clinical patterns. Emerg Infect Dis. 2002;8:160–6. DOIPubMedGoogle Scholar
- Fung CP, Chang FY, Lee SC, Hu BS, Kuo BI, Liu CY, A global emerging disease of Klebsiella pneumoniae liver abscess: is serotype K1 an important factor for complicated endophthalmitis? Gut. 2002;50:420–4. DOIPubMedGoogle Scholar
- Wang JH, Liu YC, Lee SS, Yen MY, Chen YS, Wang JH, Primary liver abscess due to Klebsiella pneumoniae in Taiwan. Clin Infect Dis. 1998;26:1434–8. DOIPubMedGoogle Scholar
- Wong TY, Chiu SI, So MK, Tsang MK, Lai ST, Tse KK, Septic metastatic endophthalmitis complicating Klebsiella pneumoniae liver abscess in a non-diabetic Chinese man. Hong Kong Med J. 2001;7:303–6.PubMedGoogle Scholar
- Chang SC, Fang CT, Hsueh PR, Chen YC, Luh KT. Klebsiella pneumoniae isolates causing liver abscess in Taiwan. Diagn Microbiol Infect Dis. 2000;37:279–84. DOIPubMedGoogle Scholar
- Lau YJ, Hu BS, Wu WL, Lin YH, Chang HY, Shi ZY. Identification of a major cluster of Klebsiella pneumoniae isolates from patients with liver abscess in Taiwan. J Clin Microbiol. 2000;38:412–4.PubMedGoogle Scholar
- Tsay RW, Siu LK, Fung CP, Chang FY. Characteristics of bacteremia between community-acquired and nosocomial Klebsiella pneumoniae infection: risk factor for mortality and the impact of capsular serotypes as a herald for community-acquired infection. Arch Intern Med. 2002;162:1021–7. DOIPubMedGoogle Scholar
- Carpenter JL. Klebsiella pulmonary infections: occurrence at one medical center and review. Rev Infect Dis. 1990;12:672–82.PubMedGoogle Scholar
- Korvick JA, Hackett AK, Yu VL, Muder RR. Klebsiella pneumonia in the modern era: clinicoradiographic correlations. South Med J. 1991;84:200–4.PubMedGoogle Scholar
- Hammond JM, Potgieter PD, Linton DM, Forder AA. Intensive care management of community-acquired Klebsiella pneumoniae. Respir Med. 1991;85:11–6. DOIPubMedGoogle Scholar
- Feldman C, Kallenbach JM, Levy H, Thorburn JR, Hurwitz MD, Koornhof HJ. Comparison of bacteraemic community-acquired lobar pneumonia due to Streptococcus pneumoniae and Klebsiella pneumoniae in an intensive care unit. Respiration. 1991;58:265–70. DOIPubMedGoogle Scholar
- Chen CW, Jong GM, Shiau JJ, Hsiue TR, Chang HY, Chuang YC, Adult bacteremic pneumonia: bacteriology and prognostic factors. J Formos Med Assoc. 1992;91:754–9.PubMedGoogle Scholar
- Feldman C, Ross S, Mahomed AG, Omar J, Smith C. The aetiology of severe community-acquired pneumonia and its impact on initial, empiric, antimicrobial chemotherapy. Respir Med. 1995;89:187–92. DOIPubMedGoogle Scholar
- Lee KH, Hui KP, Tan WC, Lim TK. Severe community-acquired pneumonia in Singapore. Singapore Med J. 1996;37:374–7.PubMedGoogle Scholar
- Garner JS, Jarvis WR, Emori TG, Horan TC, Hughes JM. CDC definitions for nosocomial infections, 1988. Am J Infect Control. 1988;16:128–40. DOIPubMedGoogle Scholar
- Palfreyman JM. Klebsiella serotyping by counter-current immunoelectrophoresis. J Hyg (Lond). 1978;81:219–25. DOIPubMedGoogle Scholar
- Orskov F, Orskov I. Immunoelectrophoretic patterns of extracts from Escherichia coli O antigen test strains O1 to O157 examinations in homologous OK sera. Further comments on the classification of Escherichia K antigens. Acta Pathol Microbiol Scand [B]. Microbiol Immunol. 1972;80:905–10.
- Hansen DS, Mestre F, Alberti S, Hernandez-Alles S, Alvarez D, Domenech-Sanchez A, Klebsiella pneumoniae lipopolysaccharide O typing: revision of prototype strains and O-group distribution among clinical isolates from different sources and countries. J Clin Microbiol. 1999;37:56–62.PubMedGoogle Scholar
- Jacobson SH, Hammarlind M, Lidefeldt KJ, Osterberg E, Tullus K, Brauner A. Incidence of aerobactin-positive Escherichia coli strains in patients with symptomatic urinary tract infection. Eur J Clin Microbiol Infect Dis. 1988;7:630–4. DOIPubMedGoogle Scholar
- Fung CP, Hu BS, Chang FY, Lee SC, Kuo BI, Ho M, A 5-year study of the seroepidemiology of Klebsiella pneumoniae: high prevalence of capsular serotype K1 in Taiwan and implication for vaccine efficacy. J Infect Dis. 2000;181:2075–9. DOIPubMedGoogle Scholar
- Podschun R, Ullmann U. Klebsiella spp. as nosocomial pathogens: epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin Microbiol Rev. 1998;11:589–603.PubMedGoogle Scholar
- Tsay RW, Siu LK, Fung CP, Chang FY. Characteristics of bacteremia between community-acquired and nosocomial Klebsiella pneumoniae infection: risk factor for mortality and the impact of capsular serotypes as a herald for community-acquired infection. Arch Intern Med. 2002;162:1021–7. DOIPubMedGoogle Scholar
- Ofek I, Kabha K, Athamna A, Frankel G, Wozniak DJ, Hasty DL, Genetic exchange of determinants for capsular polysaccharide biosynthesis between Klebsiella pneumoniae strains expressing serotypes K2 and K21a. Infect Immun. 1993;61:4208–16.PubMedGoogle Scholar
- Kabha K, Schmegner J, Keisari Y, Parolis H, Schlepper-Schaeffer J, Ofek I. SP-A enhances phagocytosis of Klebsiella by interaction with capsular polysaccharides and alveolar macrophages. Am J Physiol. 1997;272:L344–52.PubMedGoogle Scholar
- Mizuta K, Ohta M, Mori M, Hasegawa T, Nakashima I, Kato N. Virulence for mice of Klebsiella strains belonging to the O1 group: relationship to their capsular (K) types. Infect Immun. 1983;40:56–61.PubMedGoogle Scholar
- Wacharotayankun R, Arakawa Y, Ohta M, Tanaka K, Akashi T, Mori M, Enhancement of extracapsular polysaccharide synthesis in Klebsiella pneumoniae by RmpA2, which shows homology to NtrC and FixJ. Infect Immun. 1993;61:3164–74.PubMedGoogle Scholar
- Nassif X, Fournier JM, Arondel J, Sansonetti PJ. Mucoid phenotype of Klebsiella pneumoniae is a plasmid-encoded virulence factor. Infect Immun. 1989;57:546–52.PubMedGoogle Scholar
- Nassif X, Sansonetti PJ. Correlation of the virulence of Klebsiella pneumoniae K1 and K2 with the presence of a plasmid encoding aerobactin. Infect Immun. 1986;54:603–8.PubMedGoogle Scholar
Figures
Tables
Cite This Article2The International Klebsiella Study Group comprises the previously named authors plus Jose Maria Casellas, Gordon Trenholme, Joseph McCormack, Sunita Mohapatra, and Lutfiye Mulazimoglu.
Table of Contents – Volume 13, Number 7—July 2007
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Victor L. Yu, Special Pathogens Laboratory, 1401 Forbes Ave, Pittsburgh, PA 15219, USA;
Top