Volume 15, Number 10—October 2009
Dispatch
Novel Rickettsia in Ticks, Tasmania, Australia
Cite This Article
Citation for Media
Abstract
A novel rickettsia was detected in Ixodes tasmani ticks collected from Tasmanian devils. A total of 55% were positive for the citrate synthase gene by quantitative PCR. According to current criteria for rickettsia speciation, this new rickettsia qualifies as Candidatus Rickettsia tasmanensis, named after the location of its detection.
In Australia, 4 rickettsial species are known to cause disease in humans; none of these species has been identified in Tasmania. However, 3 cases of human rickettsial infections in Tasmania have been documented (1–3). Ixodes tasmani ticks are of particular interest because they are known to be vectors for other rickettsial species in Australia (4) and are also the most common tick species in Tasmania (5). In addition, because these ticks bite humans, they are candidates for rickettsial transmission in Tasmania.
Although Candidatus Rickettsia tasmanensis, a proposed new species of rickettsiae, has not been associated with human disease, the possible virulence of this rickettsia cannot be disregarded. Some initially identified rickettsiae were later found to cause disease in humans. For example, R. parkeri was discovered in 1939 (6) but was only confirmed as a human pathogen in 2004 (7). To investigate infections in Tasmania, we collected ticks from Tasmanian devils (Sacrophilus harrissi) and analyzed them for rickettsial species.
Forty-four I. tasmani ticks were collected from Tasmanian devils from various sites in Tasmania during 2005–2006; 36 were engorged females, 5 were nymphs, and 3 were males. Each tick was washed in 70% ethanol, rinsed in sterile phosphate-buffered saline, and homogenized. Homogenates were then subjected to DNA extraction by using a QIAmp DNA Blood Mini Kit (QIAGEN, Hilden, Germany). The presence of a rickettsial agent was detected by real-time PCR (8). Characterization of novel rickettsial species was achieved by comparing sequences of genes as described (9).
Amplification and sequencing of 1,096-, 3,005-, 588-, and 4,918-bp products for the citrate synthase (gltA), surface cell antigen (sca4), outer membrane protein A (ompA), and ompB genes, respectively, were conducted by using primers previously described (9). The 16S rRNA (rrs) gene was not amplified because cell culture isolation was not performed. Amplicons were cloned by using the TA Cloning Kit (Invitrogen, Carlsbad, CA, USA) and extracted by using a QuickLyse Mini Prep Kit (QIAGEN).
Big Dye sequencing was performed by using a GeneAmp PCR System 2400 thermocycler (Applied Biosystems, Foster City, CA, USA). Resulting products were analyzed at the Australian Genomic Research Facility by using an ABI Prism 3730xl DNA Analyzer (Applied Biosystems).
Sequences were assembled and edited by using the SeqMan Pro program (DNASTAR, Inc., Madison, WI, USA) and evaluated by using neighbor-joining and maximum-parsimony methods in MEGA 4 (10) and the maximum-likelihood method in PHYLIP (11). Results were confirmed by using BLAST analysis software (http://blast.ncbi.nlm.nih.gov/Blast.cgi). All sequences have been deposited in GenBank (Table).
Figure 1
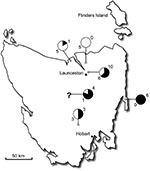
Figure 1. Map of Tasmania, Australia, showing number of positive (black) and negative (white) ticks and their locations. The question mark indicates unknown locations. A total of 55% of the ticks were positive...
Rickettsial DNA was detected in 24 (55%) of 44 I. tasmani ticks by using a gltA-specific quantitative PCR (qPCR) assay. Because most ticks were engorged females, no statistical correlation was found between the sex of ticks and presence of rickettsiae. Distribution of the ticks collected and degree of positivity are shown in Figure 1.
Sequences from I. tasmani ticks were compared with reported sequences (12). Results showed that the closest phylogenetic relative for 3 of the genes was R. raoultii strain Khabarovsk, with sequence similarities of 99.1% (1,086 bp/1,096 bp), 96.9% (570 bp/588 bp), and 97.7% (4,782 bp/4,895 bp) for the gltA, ompA, and ompB genes, respectively, and 98.1% (2,930 bp/2,988 bp) to R. japonica strain YM for the sca4 gene.
Comparison of our sequences with that of a partially sequenced rickettsia (R. tasmanensis strain T120) previously detected in an I. tasmani tick removed from a child near Underwood, Tasmania (N. Unsworth, unpub. data) found homology levels to be within the species threshold. No data on the clinical state of the child were obtained.
Sequences closely matched genes of a second partially sequenced rickettsia (Rickettsia sp. 518) from an I. tasmani tick removed from a Tasmanian devil in Tasmania by researchers at Macquarie University (Sydney, New South Wales, Australia) (13). Of the 3 partial gene sequences reported, ompB and gltA gene sequences matched to the species level with Candidatus R. tasmanensis; however, ompA gene sequences did not. Their isolate could be another new species, although it is difficult to draw conclusions with sequences of small fragments.
Figure 2

Figure 2. Phylogenetic tree showing the relationship of a 4,834-bp fragment of the outer membrane protein B gene of Candidatus Rickettsia tasmanensis (in boldface) among all validated rickettsia species. The tree was prepared...
Results of sequence analysis of the ompB gene by using the neighbor-joining algorithm are shown in Figure 2. Although all selected genes were analyzed, the ompB gene tree had the strongest bootstrap values and the largest compared fragment size.
All 44 ticks were collected from northeastern and eastern Tasmania. The number of positive samples (55%) contrasts with the small number of reported SFG rickettsial human infections in Tasmania because I. tasmani, which is known to opportunistically bite humans (5), has a high density in Tasmania. Clinical cases of infection may be missed because physicians are not aware of human rickettsial disease in Tasmania.
A recent study showed a high level of exposure to SFG rickettsia in cats and dogs near the city of Launceston, Tasmania (L. Izzard, unpub. data); in the Ravenswood area 10 of 16 tick samples were positive for SFG rickettsia by qPCR. However, the species of SFG rickettsiae could not be determined in this study because only serologic analysis was conducted. Because I. tasmani ticks are common in Tasmania and parasitize cats and dogs, Candidatus R. tasmanensis is likely to be the causative agent in some of the cases.
When gene sequences of Candidatus R. tasmanensis were compared with those of validated species (12), they did not closely match either of the 2 validated SFG rickettsia in Australia (R. australis or R. honei). Similarly, Candidatus R. tasmanensis sequences were divergent from 2 Candidatus species (Candidatus R. gravesii and Candidatus R. antechini) in Australia, which are currently being characterized. Candidatus R. tasmanensis had the highest phylogenetic similarity to R. raoultii strain Khabarovsk for 3 of 4 gene sequences. This rickettsial species was isolated in the Russian Far East (>10,000 km north of Tasmania) from a Dermacentor silvarum tick and is a known human pathogen (12). However, the similarities between the gene sequences of these 2 organisms were well below the threshold defined by Fournier et al. (9). On the basis of these results, we propose to give this Rickettsia sp. a Candidatus status and formally name it Candidatus R. tasmanensis after the location from which it was originally isolated. To validate Candidatus R. tasmanensis as a novel species, isolation and subsequent sequencing of its rrs gene are needed. Multigene sequencing of 4 other qPCR-positive I. tasmani ticks will also be useful.
Because the range of this study was limited to eastern Tasmania, I. tasmani ticks from western Tasmania and other parts of Australia should also be examined for this rickettsial agent. This analysis would help determine its true range. Testing the blood of animals infested with I. tasmani ticks for evidence of SFG rickettsial exposure may also provide data on the pathogenesis and range of this rickettsia.
Mr Izzard is a doctoral candidate at Murdoch University in Perth and works at the Australian Rickettsial Reference Laboratory in Geelong. His research interests are detection and isolation of novel rickettsial species and molecular diagnostics of rickettsial pathogens in Australia.
Acknowledgment
We thank Ian Norton, his colleagues, and Dydee Mann for collecting ticks from sites in Tasmania; and Ian Beveridge for assistance with tick speciation.
References
- Unsworth NB, Stenos J, Graves SR, Faa AG, Cox GE, Dyer JR, Flinders Island spotted fever rickettsioses caused by “marmionii” strain Rickettsia honei, eastern Australia. Emerg Infect Dis. 2007;13:566–73. DOIPubMedGoogle Scholar
- Chin RH, Jennens ID. Rickettsial spotted fever in Tasmania. Med J Aust. 1995;162:669.PubMedGoogle Scholar
- Watts MR, Benn RA, Hudson BJ, Graves S. A case of prolonged fatigue following an acute rickettsial infection. QJM. 2008;101:591–3. DOIPubMedGoogle Scholar
- Sexton DJ, Dwyer B, Kemp R, Graves S. Spotted fever group rickettsial infections in Australia. Rev Infect Dis. 1991;13:876–86.PubMedGoogle Scholar
- Roberts FH. Australian ticks. Melbourne: Commonwealth Scientific and Industrial Research Organisation; 1970.
- Parker R, Kohls G, Cox G, Davis G. Observations on an infectious agent from Amblyomma maculatum. Public Health Rep. 1939;54:1482–4.
- Paddock CD, Sumner JW, Comer JA, Zaki SR, Goldsmith CS, Goddard J, Rickettsia parkeri: a newly recognized cause of spotted fever rickettsiosis in the United States. Clin Infect Dis. 2004;38:805–11. DOIPubMedGoogle Scholar
- Stenos J, Graves S, Unsworth N. A highly sensitive and specific real-time PCR assay for the detection of spotted fever and typhus group rickettsiae. Am J Trop Med Hyg. 2005;73:1083–5.PubMedGoogle Scholar
- Fournier PE, Dumler JS, Greub G, Zhang J, Wu Y, Raoult D. Gene sequence-based criteria for identification of new Rickettsia isolates and description of Rickettsia heilonjiangensis sp. nov. J Clin Microbiol. 2003;41:5456–65. DOIPubMedGoogle Scholar
- Tamura K, Dudley J, Nei M, Kumar S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol Biol Evol. 2007;24:1596–9. DOIPubMedGoogle Scholar
- Felsenstein J. PHYLIP: Phylogeny Inference Package (Version 3.2). Cladistics. 1989;5:164–6.
- Mediannikov O, Matsumoto K, Samoylenko I, Drancourt M, Roux V, Rydkina E, Rickettsia raoultii sp. nov., a spotted fever group rickettsia associated with Dermacentor ticks in Europe and Russia. Int J Syst Evol Microbiol. 2008;58:1635–9. DOIPubMedGoogle Scholar
- Vilcins IM, Old JM, Deane E. Detection of a Hepatozoon and spotted fever group Rickettsia species in the common marsupial tick (Ixodes tasmani) collected from wild Tasmanian devils (Sarcophilus harrisii), Tasmania. Vet Parasitol. 2009;162:23–31. DOIPubMedGoogle Scholar
Figures
Table
Cite This ArticleTable of Contents – Volume 15, Number 10—October 2009
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Leonard Izzard, Australian Rickettsial Reference Laboratory, The Geelong Hospital, Barwon Health, PO Box 281, Geelong, Victoria, 3220 Australia
Top