Volume 15, Number 2—February 2009
Dispatch
European Bat Lyssavirus Transmission among Cats, Europe
Cite This Article
Citation for Media
Abstract
We identified 2 cases of European bat lyssavirus subtype 1 transmission to domestic carnivores (cats) in France. Bat-to-cat transmission is suspected. Low amounts of virus antigen in cat brain made diagnosis difficult.
Most countries in western Europe are currently free of rabies in terrestrial mammals, as was the case in France during 2001–2008 (1). However, rabies still remains a public health problem in these countries because of natural circulation of bat-specific viruses (order Mononegavirales, family Rhabdoviridae, genus Lyssavirus) such as European bat lyssaviruses (EBLVs). These viruses are divided into genotypes 5 (EBLV-1) and 6 (EBLV-2); the first genotype is subdivided into subtypes a and b (2). Knowledge of the prevalence and epidemiology of EBLV is limited (2–5). To date, natural transmission of EBLV-1 has been reported in a limited number of terrestrial mammals, including 5 sheep in Denmark (6) and 1 stone marten in Germany (7) (Table 1). Since 1985, only 3 human deaths from EBLVs have been confirmed (3) (Table 1). We describe 2 documented cases of spillover transmission of EBLV in domestic carnivores (cats, Felis domesticus) in Europe.
In November 2003, a 6-month-old female stray cat (cat no. 1) was found ill in a public garden in Vannes (Morbihan District) in western France and taken to a veterinary clinic. This animal had convulsions and moderate dehydration and was emaciated. It was infected by feline immunodeficiency virus, which was compatible with the clinical symptoms. The veterinarian was bitten while providing veterinary care to the cat. After a few days, the cat recovered and was impounded for veterinary surveillance. It died suddenly the following night. No information about potential contact with bats was available.
On November 8, 2007, an 18-month-old female cat (cat no. 2) was taken by its owner to a veterinarian in Fontenay-le-Comte (Vendée District) in western France because of abnormal behavior. The owner reported having been bitten by the cat. The next day, the cat showed severe central neurologic disorders and aggressive behavior. It died during the next night. Its outdoor access appeared to have been restricted. Two months later, the carcass of a bat (Eptesicus serotinus) was recovered in the same area of Fontenay-le-Comte and submitted for rabies testing.
Recommended techniques for rabies diagnosis were used for all animals (8). For cat no. 1, results of a repeated direct immunofluorescence antibody test (FAT) with a polyclonal antirabies conjugate (Bio-Rad, Marnes-la-Coquette, France) performed on different cortex and spinal bulb smears were negative. Viral isolation by using a rabies tissue culture infection test (RTCIT) was also unsuccessful, as was attempted isolation of virus by using a mouse inoculation test (MIT) (Table 1). The only test routinely used that gave a positive result was an antigen-capture ELISA (WELYSSA) for lyssavirus antigen (9). The presence of EBLV RNA (03011FRA) was determined by reverse transcription–PCR (RT-PCR) targeting short viral gene regions (5).
Lyssavirus antigens were repeatedly detected by FAT in different areas of the brain of cat no. 2. Viral isolation by using RTCIT was positive only after the second cell culture passage. Results for isolation of EBLV (07240FRA) by MIT were positive. Lyssavirus antigen detection by WELYSSA was variable, depending on the part of the brain tested. Viral RNA was detected by RT-PCR (Table 1). The bat was positive for EBLV by FAT, RTCIT (08120FRA), MIT, and RT-PCR.
Figure 1

Figure 1. Phylogenetic tree comparing nucleotide sequences of A) nucleoprotein (372 nt, position 63 from the translation initiation site) and B) glycoprotein (547 nt, position 640 from the translation initiation site) genes of...
Nucleotide sequencing and phylogenetic analysis identified isolate 03011FRA as EBLV-1b and isolates 08120FRA and 07240FRA as EBLV-1a (Figure 1). Sequencing of the complete genome (10) of the 2 EBLV-1a isolates showed a high percentage of homology (Table 2).
After identification of these 2 cases of spillover transmission of EBLV-1 to domestic cats, postexposure prophylaxis measures were implemented. The veterinarian who was bitten by cat no. 1 received a booster rabies vaccination, and 15 persons exposed to cat no. 2 during the 2-week critical period before its death received appropriate postexposure treatment on the basis of national and international recommendations of the World Health Organization (8,11). Two family dogs potentially exposed to cat no. 2 and previously vaccinated received a booster vaccination. Cross-neutralization data obtained with human serum samples and in rodent models suggest that preexposure and postexposure treatments for rabies are effective against EBLV-1.
Control measures were implemented to prevent potential further contaminations, although cats represent naturally dead-end host for rabies (and for lyssavirus), thereby limiting any risk for transmission to other mammals. City authorities conducted a census of all domestic animals in the neighborhood where cat no. 2 lived. All cats, dogs, and ferrets were identified by microchips and kept under veterinary surveillance. Dogs had to be leashed and cats kept indoors during the next 2 months.
We report 2 documented cases of natural infection of domestic cats by EBLV-1 lyssaviruses presently circulating in European bats (2–5). Our study demonstrates that subtypes EBLV-1a and EBLV-1b can cross the species barrier, although cat no. 1 probably died of feline leukemia. However, cat no. 2 died with neurologic signs compatible with rabies and was positive for EBLV by FAT, the reference technique. Direct transmission of EBLV-1 from bats to cats seems the most realistic explanation for these cases because cats prey on bats and have numerous contacts with them (3,12,13). Identification of a highly homologous EBLV-1a isolate from a rabid bat found in the same location as cat no. 2 supports this hypothesis.
Difficulties in EBLV-1 detection in the brain of these 2 cases of spillover transmission are reminiscent of transmission previously reported (6,7) (Table 1), potentially caused by a low amount of virus antigen in the brain. This finding further underlines the importance of using different techniques to diagnose rabies caused by EBLVs (9). This issue raises questions about the true incidence of these viruses among cats.
Figure 2
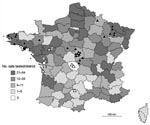
Figure 2. Distribution of cats analyzed during 2004–2007 and of bats found positive for European bat lyssavirus (EBLV) in France during 1989–2007. Distribution of 1,506 cats tested during 2004–2007 by direct immunofluorescence antibody...
At the Institut Pasteur during 1997–2007, a total of 6,097 cats suspected of having rabies and originating from all districts in France showed negative results for rabies by 2 recommended techniques (FAT and RTCIT). Among them, all animals tested since 2004 (1,506 cats), except cats no. 1 and no. 2, were also negative for rabies by WELYSSA (Figure 2), which suggests that transmission of EBLVs from bats to cats, although possible, is rare. Furthermore, terrestrial mammals seem to represent dead-end hosts for EBLVs, as suggested by results of experimental EBLV inoculations in several mammals such as cats, dogs, ferrets, mice, red foxes, or sheep (6,7,14). These animals are susceptible to infection with EBLVs but seem unlikely to actively transmit EBLVs to a new host.
Comparative analysis of the full-length genomic sequence of the EBLV-1a from cat no. 2 isolate 07240FRA with bat isolate 08120FRA and with another bat (E. serotinus) isolate (03002FRA) collected in 2003 ≈100 km from Fontenay-le-Comte showed high similarity (Table 2). This finding indicates that heterologous passage of EBLV-1a in a cat did not select mutants. Similarly, the lower similarity observed with a genomic sequence collected from a bat RV9 (15) (E. serotinus) in 1968 in Germany also indicates that EBLV-1 evolution is shaped by slow genetic drift (2).
No secondary cases originating from cat no. 1 and cat no. 2 were reported (after 6 months of follow-up for cat no. 2). However, improving surveillance and raising awareness to better understand the epidemiology of lyssaviruses are necessary. Persons bitten by bats or by any carnivorous animal are advised to wash wounds with water and soap and to seek medical attention (8,11).
Dr Dacheux works in the UPRE Lyssavirus Dynamics and Host Adaptation unit at the Institut Pasteur in Paris and at the National Reference Centre for Rabies in Paris. His research interest is rabies, particularly its surveillance, development of new diagnostic tools, and physiopathologic studies in humans and animals.
Acknowledgments
We thank P. Cozette, R. Lavenir, L. Audry, and M. Tichit for expert technical assistance; Lisa King for critical review of the manuscript; Zélie Lardon for help with Arctique software; Chiraz Talbi for help with Bayesian Evolutionary Analysis Sampling Trees software; and Evelyne Picard-Meyer for permission to use bat data from the Agence Française de Sécurité Sanitaire des Aliments–Malzeville.
This study was supported by the European Commission through the “VIZIER” Integrated Project (LSHG-CT-2004-511966), a grant from the Genomic Platform Ile-de-France “Analyse de la Diversité Génétique des Rhabdoviridae,” 2005–2006, and the Institut de Veille Sanitaire, Saint-Maurice, France.
References
- Bourhy H, Dacheux L, Strady C, Mailles A. Rabies in Europe in 2005. Euro Surveill. 2005;10:213–6.PubMedGoogle Scholar
- Davis PL, Holmes EC, Larrous F, Van der Poel WH, Tjørnehøj K, Alonso WJ, Phylogeography, population dynamics, and molecular evolution of European bat lyssaviruses. J Virol. 2005;79:10487–97. DOIPubMedGoogle Scholar
- Harris SL, Brookes SM, Jones G, Hutson AM, Racey PA, Aegerter J, European bat lyssaviruses: distribution, prevalence and implications for conservation. Biol Conserv. 2006;131:193–210. DOIGoogle Scholar
- Vos A, Kaipf I, Denzinger A, Fooks AR, Johnson N, Müller T. European bat lyssaviruses: an ecological enigma. Acta Chiropt. 2007;9:283–96. DOIGoogle Scholar
- Amengual B, Bourhy H, López-Roig M, Serra-Cobo J. Temporal dynamics of European bat lyssavirus type 1 and survival of Myotis myotis bats in natural colonies. PLoS One. 2007;2:e566. DOIPubMedGoogle Scholar
- Tjørnehøj K, Fooks AR, Agerholm JS, Rønsholt L. Natural and experimental infection of sheep with European bat lyssavirus type-1 of Danish bat origin. J Comp Pathol. 2006;134:190–201. DOIPubMedGoogle Scholar
- Müller T, Cox J, Peter W, Schäfer R, Johnson N, McElhinney LM, Spill-over of European bat lyssavirus type 1 into a stone marten (Martes foina) in Germany. J Vet Med B Infect Dis Vet Public Health. 2004;51:49–54. DOIPubMedGoogle Scholar
- World Health Organization. WHO expert consultation on rabies. World Health Organ Tech Rep Ser. 2005;931:1–88.PubMedGoogle Scholar
- Xu G, Weber P, Hu Q, Xue H, Audry L, Li C, A simple sandwich ELISA (WELYSSA) for the detection of lyssavirus nucleocapsid in rabies suspected specimens using mouse monoclonal antibodies. Biologicals. 2007;35:297–302. DOIPubMedGoogle Scholar
- Delmas O, Holmes EC, Talbi C, Larrous F, Dacheux L, Bouchier C, Genomic diversity and evolution of the lyssaviruses. PLoS One. 2008;3:e2057. DOIPubMedGoogle Scholar
- Conseil Supérieur d’Hygiène Publique de France (CSHPF). Recommandations pour limiter l’exposition du public aux virus de la rage des chauves-souris. Bulletin Epidémiologique Hebdomadaire. 2001;39:193.
- Takumi K, Lina PH, van der Poel WH, Kramps JA, van der Giessen JW. Public health risk analysis of European bat lyssavirus infection in The Netherlands. Epidemiol Infect. 2008;21:1–7. DOIGoogle Scholar
- Tjørnehøj K, Rønsholt L, Fooks AR. Antibodies to EBLV-1 in a domestic cat in Denmark. Vet Rec. 2004;155:571–2.PubMedGoogle Scholar
- Brookes SM, Klopfleisch R, Müller T, Healy DM, Teifke JP, Lange E, Susceptibility of sheep to European bat lyssavirus type-1 and -2 infection: a clinical pathogenesis study. Vet Microbiol. 2007;125:210–23. DOIPubMedGoogle Scholar
- Marston DA, McElhinney LM, Johnson N, Müller T, Conzelmann KK, Tordo N, Comparative analysis of the full genome sequence of European bat lyssavirus type 1 and type 2 with other lyssaviruses and evidence for a conserved transcription termination and polyadenylation motif in the G-L 3′ non-translated region. J Gen Virol. 2007;88:1302–14. DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 15, Number 2—February 2009
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Laurent Dacheux, UPRE Dynamique des Lyssavirus et Adaptation à l’Hôte, Centre National de Réference pour la Rage, Institut Pasteur, 25 Rue du Docteur Roux, 75724 Paris Cedex 15, France;
Top