Volume 16, Number 1—January 2010
Dispatch
Seagulls and Beaches as Reservoirs for Multidrug-Resistant Escherichia coli
Cite This Article
Citation for Media
Abstract
A variety of extended-spectrum β-lactamase–producing Escherichia coli isolates, with a high rate of cefotaximase-15 resistance, were identified in seagull feces from Porto, Portugal, beaches. Beaches may therefore present a risk to public health because of the potential pathogen-spreading capacity of migratory birds.
The Porto coastline in Portugal, including downtown Porto, has a large seagull population (Larus fuscus, the lesser black-backed gull, and L. cachinnans, the yellow-legged gull). Seagulls have been reported as a possible reservoir for multidrug-resistant bacteria (1). During the past decade, extended-spectrum β-lactamases (ESBL) with growing epidemiologic importance, the cefotaximase (CTX-M) enzymes, have been reported worldwide in Enterobacteriaceae from humans (2); they are found mostly in Escherichia coli. Reasons for the emergence of CTX-M enzymes in human isolates remain unknown. In addition, CTX-M–positive E. coli have been identified in poultry (3), other farm animals (4), and wild animals (birds of prey, foxes [5]). E. coli strains can be classified into 4 phylogenetic groups (A, B1, B2, and D). The virulent extraintestinal isolates belong mostly to group B2 and, to a lesser extent, group D, whereas most commensal strains belong to groups A and B1. The objective of the study was to evaluate the spread and types of ESBL-positive E. coli in feces recovered from wild birds on the beaches of Porto.
During December 2007 through April 2008, wild seagull (L. fuscus, L. cachinnans) feces were collected from the Matosinhos and Leça da Palmeira beaches (Porto, Portugal) (20 samples every 2 weeks) using a sterile spatula. Care was taken during sampling to avoid collection of beach sediment. Samples were placed in sterile tubes and processed. Samples were precultured in buffered peptone water (Oxoid, Basingstoke, UK) at a dilution of 1/10 wt/vol and incubated at 37°C. Cultures were injected by streaking 10 µL of the suspensions onto Tergitol BCIG agar (Biokar Diagnostics, Beauvais, France). Another suspension was made in buffered peptone water supplemented with cefotaxime (CTX) at 2 µg/mL and then streaked in a Tergitol BCIG plate supplemented with CTX at 2 µg/mL. The plates were incubated at 37°C overnight. We identified E. coli isolates by using the API20E system (bioMérieux, Balmes-les-Grottes, France).
Susceptibility testing was performed by disk diffusion assay (Sanofi-Diagnostic Pasteur, Marnes-la-Coquette, France), as previously described (3). MICs were determined by Etest (AB BIODISK; Solna, Sweden) on Mueller-Hinton agar plates at 37°C (8). ESBL was detected with a synergy test using disks containing CTX, ceftazidime, and ticarcillin-clavulanic acid (6).
Clonal diversity was assessed by pulsed-field gel electrophoresis (PFGE) as described (3). Genomic DNA was extracted in situ by treatment with lysozyme (1 mg/mL; Sigma, Saint-Quentin Fallavier, France) and proteinase K (0.5 mg/mL; Sigma) and then restricted with endonuclease XbaI (GE Healthcare, Aulnay-sous-Bois, France). We separated resulting fragments by using 1% PFGE-grade agarose gel (Bio-Rad, Hercules, CA, USA) in a CHEF-DR II System (Bio-Rad), with the following protocol: 6 volts/cm, 4–12 s pulse time for 12 h, followed by 15–36 s pulse time for 12 h in 0.5% tris-borate-EDTA buffer at 14°C.
Detection of blaCTX-M genes was carried out by PCR (3). We sequenced the purified PCR products on both strands by using an Applied Biosystems sequencer (ABI 377; Foster City, CA, USA) and analyzed these sequences in the BLAST database (www.ncbi.nlm.nih.gov/blast/Blast.cgi). Phylogenetic grouping of E. coli isolates was determined by PCR (7) for assignation of phylogenetic groups: group B2, chuA+, yjaA+; group D, chuA+, yjaA–; group B1, chuA–, TspE4C2+; and group A, chuA–, TspE4C2–. Analysis of plasmid content was performed for the blaCTX-M-like positive isolates by using the Kieser technique (3). The multilocus sequence typing (MLST) of E. coli isolates was determined by sequencing 7 essential genes (adk, fumC, icd, purA, gyrB, recA, and mdh) as described (8), followed by an analysis on the E. coli MLST website (http://mlst.ucc.ie/mlst/dbs/Ecoli) except for mdh, icd, and recA as performed in another study (9).
We obtained 139 E. coli isolates, of which 45 (32%) displayed an ESBL phenotype. Forty-four (98%) of the 45 ESBL producers carried a blaCTX-M gene; 1 isolate possessed a blaTEM-52 gene. PCR and sequencing identified the CTX-M ESBL determinants as follows: 8 (18%) were CTX-M-1; 4 (9%), CTX-M-9; 17 (39%), CTX-M-15; and 15 (34%), CTX-M-32.
PGFE analysis showed a high diversity of genotypes: 8 clones for 8 CTX-M-1–positive isolates, 4 clones for 4 CTX-M-9–positive isolates, 13 clones for 17 CTX-M-15–positive isolates, and 14 clones for 15 CTX-M-32–positive isolates (data not shown). A total of 37% of ESBL producers belonged to virulent extraintestinal groups B2 and D. Of these isolates, 41% expressed CTX-M-15 and 47% expressed CTX-M-32.
MLST identified 25 different types among the 45 E. coli isolates. The most commonly identified genotypes were ST1284 (4 isolates), ST131 (4), and ST224 (3). Isolates belonging to ST453, ST86, ST205, ST359, ST165, ST69, ST1152, ST405, ST559, ST1163, ST10, ST58, ST156, ST155, ST10, ST297, ST43, ST58, and ST156 were also identified. Different genotypes carrying the same ESBL determinant were identified; conversely, different ESBL determinants were found among the same genotype. In particular, isolates belonging to the ST131 genotype widely reported among human CTX-M-15–positive E. coli producers (9,10) harbored either blaCTX-M-32 (2 isolates), blaCTX-M-1 (1), or blaCTX-M-15 (1). Coresistances of the 45 ESBL-positive isolates were as follows: 90% were resistant to tetracycline; 60%, to trimethoprim/sulfamethoxazole; 55%, to nalidixic acid; 50%, to ciprofloxacin; 23%, to gentamicin; and 4%, to chloramphenicol.
Figure
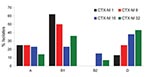
Figure. Distribution of isolates positive for cefotaximase, by phylogenetics group. Vertical axis indicates the percentage for each CTX-M determinant.
Identification of the phylogroups of the different CTX-M producers showed that most belonged to phylogroup B1, and a notable number of isolates belonged to phylogroup D (Figure). In contrast, few isolates belonged to the highly virulent phylogroup B2. Whereas CTX-M-1 producers mostly belonged to phylogroup B1, the CTX-M-15 producers were well distributed among the 4 phylogroups (Figure). Notably, plasmid analysis identified a sizable diversity of plasmid sizes, including E. coli isolates producing the same CTX-M type (data not shown).
CTX-M type β-lactamases are ESBLs of increasing prevalence (2). Infections with multidrug-resistant bacilli occur not only in hospitals but also in the community (11). Previous studies have reported multidrug-resistance in wild birds (12,13). Our study provides additional clues that wild seagulls are carriers of ESBL-producing E. coli, although at a lower rate than previously reported (1). In that study, ESBL determinants were TEM-52, CTX-M-1, CTX-M-14a, and CTX-M-32 (1). Another study (focused on poultry) (3) reported that the main identified CTX-M determinants were of the CTX-M-1 group (CTX-M-1, CTX-M-15, CTX-M-32), as we also found.
We report that CTX-M-15 was the main CTX-M type identified among birds residing mostly on beaches of downtown Porto, which agrees with CTX-M-15 being the most prevalent ESBL in E. coli in Porto-hospitalized patients (14). This observation differs from those of a study reporting mostly TEM-52 producers in wild animals in a reserve near Porto, where indirect contacts with humans are less likely (5).
Previous studies have reported the association of E. coli isolates of groups B2 and D with extraintestinal infections (15). Our report shows that 37% of all ESBL isolates belong to B2 or D phylogroup, a higher rate than previously reported (27% of all ESBL) (1). This finding could be a matter of concern for human health. However, we showed that the ST131 type known to be frequently isolated in humans and frequently associated with CTX-M-15 production was quite rare (9%). In addition, those ST131 strains were found to harbor diverse CTX-M determinants. In fact, the frequent identification of CTX-M producers here was related neither to the dissemination of a single clone nor to that of a single plasmid.
Our report suggests that beaches may play a major role in dissemination of resistance determinants and may be a source of the CTX-M-15–related community-acquired infections. Migratory birds, such as seagulls crossing an extensive portion of the European coastline between Portugal and Scandinavia, may be reservoirs for these emerging resistance determinants.
Dr Simões is a veterinary student at Instituto de Ciências Biomédicas Abel Salazar, Porto, and completed this study as the final professional training period of his studies curriculum. His research interests include zoonotic diseases.
Acknowledgments
We thank all the members of Instituto de Ciências Biomédicas Abel Salazar Departamento de Produção Aquática and all who helped and shared knowledge as part of this study.
This work was financed by a grant from the Ministère de la Recherche, Université Paris XI, Paris, France; by grants from the European Community (DRESP2, LSHM-CT-2005-01705 and TROCAR HEALTH-F3-2008-223031); and by the Institut national de la santé de la recherche médical, France.
References
- Poeta P, Radhouani H, Igrejas G, Goncalves A, Carvalho C, Rodrigues J, Seagulls of the Berlengas natural reserve of Portugal as carriers of fecal Escherichia coli harboring CTX-M and TEM extended-spectrum β-lactamases. Appl Environ Microbiol. 2008;74:7439–41. DOIPubMedGoogle Scholar
- Livermore DM, Canton R, Gniadkowski M, Nordmann P, Rossolini GM, Arlet G, CTX-M: changing the face of ESBLs in Europe. J Antimicrob Chemother. 2007;59:165–74. DOIPubMedGoogle Scholar
- Girlich D, Poirel L, Carattoli A, Kempf I, Lartigue MF, Bertini A, Extended-spectrum β-lactamase CTX-M-1 in Escherichia coli isolates from healthy poultry in France. Appl Environ Microbiol. 2007;73:4681–5. DOIPubMedGoogle Scholar
- Meunier D, Jouy E, Lazizzera C, Kobisch M, Madec JY. CTX-M-1– and CTX-M-15–type β-lactamases in clinical Escherichia coli isolates recovered from food-producing animals in France. Int J Antimicrob Agents. 2006;28:402–7. DOIPubMedGoogle Scholar
- Costa D, Poeta P, Sáenz Y, Vinué L, Rojo-Bezares B, Jouini A, Detection of Escherichia coli harbouring extended-spectrum β-lactamases of the CTX-M, TEM and SHV classes in faecal samples of wild animals in Portugal. J Antimicrob Chemother. 2006;58:1311–2. DOIPubMedGoogle Scholar
- Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing; eighteenth informational supplement. M100–S18. Wayne (PA): The Institute; 2008.
- Clermont O, Bonacorsi S, Bingen E. Rapid and simple determination of Escherichia coli phylogenetic group. Appl Environ Microbiol. 2000;66:4555–8. DOIPubMedGoogle Scholar
- Tartof SY, Solberg OD, Manges AR, Riley LW. Analysis of a uropathogenic Escherichia coli clonal group by multilocus sequence typing. J Clin Microbiol. 2005;43:5860–4. DOIPubMedGoogle Scholar
- Nicolas-Chanoine MH, Blanco J, Leflon-Guibout V, Demarty R, Alonso MP, Caniça MM, Intercontinental emergence of Escherichia coli clone O25:H4-ST131 producing CTX-M-15. J Antimicrob Chemother. 2008;61:273–81. DOIPubMedGoogle Scholar
- Lau SH, Kaufmann ME, Livermore DM, Woodford N, Willshaw GA, Cheasty T, UK epidemic Escherichia coli strains A-E, with CTX-M-15 β-lactamase, all belong to the international O25:H4-ST131 clone. J Antimicrob Chemother. 2008;62:1241–4. DOIPubMedGoogle Scholar
- Mendonça N, Leitão J, Manageiro V, Ferreira E, Caniça M. Spread of extended-spectrum β-lactamase CTX-M-producing Escherichia coli clinical isolates in community and nosocomial environments in Portugal. Antimicrob Agents Chemother. 2007;51:1946–55. DOIPubMedGoogle Scholar
- Sjölund M, Bonnedahl J, Hernandez J, Bengtsson S, Cederbrant G, Pinhassi J, Dissemination of multidrug-resistant bacteria into the Arctic. Emerg Infect Dis. 2008;14:70–2. DOIPubMedGoogle Scholar
- Cole D, Drum DJV, Stallknecht DE, White DG, Lee MD, Ayers S, Free-living Canada geese and antimicrobial resistance. Emerg Infect Dis. 2005;11:935–8.PubMedGoogle Scholar
- Machado E, Coque TM, Cantón R, Baquero F, Sousa JC, Peixe L. Dissemination of Enterobacteriaceae harboring blaCTX-M-15, blaOXA-1, blaTEM-1 and aac(6′)-Ib-cr gene in Portugal. Antimicrob Agents Chemother. 2006;50:3220–1. DOIPubMedGoogle Scholar
- Picard B, Garcia JS, Gouriou S, Duriez P, Brahimi N, Bingen E, The link between phylogeny and virulence in Escherichia coli extraintestinal infection. Infect Immun. 1999;67:546–53.PubMedGoogle Scholar
Figure
Cite This ArticleTable of Contents – Volume 16, Number 1—January 2010
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Patrice Nordmann, Hopital de Bicetre, Service de Bacteriologie-Virologie, 78, rue du General-Leclerc Le Kremlin, Bicetre 94275, France
Top