Volume 16, Number 8—August 2010
Research
White-Nose Syndrome Fungus (Geomyces destructans) in Bats, Europe
Cite This Article
Citation for Media
Abstract
White-nose syndrome is an emerging disease in North America that has caused substantial declines in hibernating bats. A recently identified fungus (Geomyces destructans) causes skin lesions that are characteristic of this disease. Typical signs of this infection were not observed in bats in North America before white-nose syndrome was detected. However, unconfirmed reports from Europe indicated white fungal growth on hibernating bats without associated deaths. To investigate these differences, hibernating bats were sampled in Germany, Switzerland, and Hungary to determine whether G. destructans is present in Europe. Microscopic observations, fungal culture, and genetic analyses of 43 samples from 23 bats indicated that 21 bats of 5 species in 3 countries were colonized by G. destructans. We hypothesize that G. destructans is present throughout Europe and that bats in Europe may be more immunologically or behaviorally resistant to G. destructans than their congeners in North America because they potentially coevolved with the fungus.
White-nose syndrome (WNS) is a recently emerged wildlife disease in North America, which in 4 years has resulted in unprecedented deaths of hibernating bats in the northeastern United States (1–3), and is a widespread epizootic disease among bats. Although we have searched the literature describing observations of hibernating bats, we have been unable to find any similar historical accounts of white fungus growing on live hibernating bats in North America before the recent emergence of WNS.
In North America, WNS is known to affect 6 species of bats that use hibernation as their winter survival strategy: the big brown bat (Eptesicus fuscus), the eastern small-footed bat (Myotis leibii), the little brown bat (M. lucifugus), the northern long-eared bat (M. septentrionalis), the tricolored bat (Perimyotis subflavus), and the Indiana bat (M. sodalis) (1,3,4). Since its detection in February 2006 in a popular tourist cave near Albany, New York, USA, WNS has spread >1,300 km into Connecticut, Massachusetts, New Hampshire, New Jersey, Pennsylvania, Tennessee, Vermont, Virginia, and West Virginia in the United States and the provinces of Ontario and Quebec in Canada (1,3,5) in a pattern suggesting the spread of an infectious agent.
A recently discovered psychrophilic (cold-loving) fungus, Geomyces destructans (6), has consistently been isolated from bats that meet the pathologic criteria for WNS, including colonization of skin by fungal hyphae causing characteristic epidermal erosions and ulcers that can progress to invasion of underlying connective tissue (2,7). G. destructans is identified by its distinctive asymmetrically curved conidia and has a unique taxonomic position among other Geomyces spp. described to date (6). Its closest genetic relative is G. pannorum, a ubiquitous psychrophilic, keratinolytic fungus that has been isolated from a variety of sources and geographic regions, including soil and the fur of wild mammals in France (8), floors of trains and ferryboats in Italy (9), boreal forests in Canada (10), and environmental samples from Arctic regions (11,12). G. pannorum var. pannorum has been reported as an unusual dermatophyte infecting fingernails and superficial skin of humans who have a history of close contact with soil and dust (13,14). However, G. destructans differs from other common soil fungi of North America in its ability to invade the living tissues of hibernating bats.
After WNS was described in North America (1), reports dating back to the early 1980s (15) described repeated observations of white fungal growth on muzzles of hibernating bats in Germany. However, these bats lacked the characteristics of WNS such as associated deaths. Moreover, fungus was not identified. In response to WNS in North America, researchers in Europe initiated a surveillance effort during the winter of 2008–09 for WNS-like fungal infections among hibernating populations of bats in Europe. G. destructans in Europe was previously reported in 1 hibernating bat that was sampled in France during March 2009 (16).
In this report, we describe results of a more extensive effort by scientists from 4 countries in Europe (Germany, United Kingdom, Hungary, and Switzerland) to obtain and analyze samples from hibernating bats with white patches on their faces or wing membranes. Our objectives were to identify the fungus colonizing such affected hibernating bats in Europe and to clarify its geographic distribution over a broad area of Europe.
Figure 1

Figure 1. A) Greater mouse-eared bat (Myotis myotis) with white fungal growth around its muzzle, ears, and wing membranes (photograph provided by Tamás Görföl). B) Scanning electron micrograph of a bat hair colonized...
During ongoing annual population surveys of caves and mines conducted by national nongovernmental organizations, hibernating bats with obvious fungal growth on their bodies (Figure 1, panel A) were opportunistically sampled in Germany, Switzerland, and Hungary; samples were also obtained from 2 dead bats from the same hibernaculum in the United Kingdom. Approximately 366 hibernacula were visited during mid-February–mid-April 2009: 336 in Germany, 20 in Hungary, and 10 in Switzerland. Two to 214 hibernating animals were observed at each site, with the exception of 2 sites in Germany, which harbored 2,000–7,000 animals at each site.
Samples were collected from live bats by using 2 methods. Touch imprints were obtained by holding adhesive tape against affected areas of skin or fur, or fur clippings were obtained from affected areas of bat muzzles. All species of bats in Europe are strictly protected under the Flora, Fauna, Habitat Guidelines of the European Union (92/43/EEC) (http://ec.europa.eu/environment/nature/legislation/habitatsdirective/index_en.htm) and The Agreement on the Conservation of Populations of European Bats (www.eurobats.org). We did not have permission to invasively sample or kill individual animals for histologic analysis to confirm skin infection by G. destructans (7). Samples were shipped to the Leibniz Institute of Zoo and Wildlife Research (IZW), Berlin, Germany, for further investigations.
Twenty adhesive tape samples were first screened by using light microscopy, and 21 hair samples were examined by using scanning electron microscopy for conidia characteristic of G. destructans (Figure 1, panel B). Two of the submitted samples (2 greater horseshoe bats from the United Kingdom) consisted of entire bat carcasses. Although the carcasses were examined externally for fungal growth on muzzle skin and hair, specimens were too decomposed to conduct internal pathologic examinations. Tape or hair samples from all bats were further investigated by using direct PCR amplification of fungal rRNA gene internal transcribed spacer (ITS) region DNA (ITS1, 5.8S, and ITS2). Total nucleic acids were extracted from culture, tape, or hair samples by using PrepMan Ultra Reagent (Applied Biosystems, Darmstadt, Germany) following the manufacturer’s instructions.
The rRNA gene ITS region DNA was amplified by using PCR with primers ITS4 and ITS5 (17) and GoTaq DNA polymerase (Promega, Madison, WI, USA). Cycling parameters were an initial 2-min denaturation at 98°C; followed by 30 cycles of denaturation at 98°C for 10 s, annealing at 50°C for 30 s, and extension at 72°C for 1 min; and a final extension at 72°C for 7 min. For fungal isolates, rRNA gene small subunit (SSU) DNA was amplified by using PCR with primers nu-SSU-0021–5′ (17) and nu-SSU-1750–3′ (18) as above, except the extension time was increased to 2 min. Sequencing primers were PCR primers; nu-SSU-0402–5′ (18), nu-SSU-1150–5′ (17), nu-SSU-0497–3′ (18), and nu-SSU-1184–3′ (19) were added for SSU. PCR products were sent to the Robert Koch Institute, Berlin, Germany, for direct sequencing.
Culture analyses of samples were performed at Munich University Hospital and IZW. After examining tape impressions by using light microscopy, we identified small areas with fungal conidia characteristic of G. destructans and excised them with a sterile scalpel blade. Half of the excised material was used for PCR; the remaining sample and samples of individual hairs with microscopic indication of G. destructans were immediately placed onto Sabouraud dextrose agar plates containing gentamicin and chloramphenicol and incubated at 4°C and 8°C. G. destructans isolates obtained during this study are maintained at IZW.
We obtained and analyzed samples from live bats with obvious fungal growth on their bodies found between mid-February and the end of March 2009 at 11 sites (8 in Germany, 1 in Hungary, and 2 in Switzerland). Samples were also obtained from an additional bat in Germany in February 2008 and from 2 dead bats from a site in the United Kingdom in March 2009 (Tables 1, 2) All 12 hibernacula sampled contained 1–5 animals that exhibited obvious fungal growth. Forty-three samples were obtained from these 12 hibernacula and represented 23 adult bats of 6 species: 1 Brandt bat (M. brandtii), 3 pond bats (M. dasycneme), 1 Daubenton bat (M. daubentonii), 1 lesser mouse-eared bat (M. oxygnathus), 15 greater mouse-eared bats (M. myotis), and 2 greater horseshoe bats (Rhinolophus ferrumequinum).
Figure 2
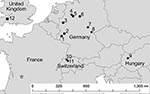
Figure 2. Locations in Europe of bats positive for Geomyces destructans by PCR alone (circles) or by PCR and culture (solid stars) and bats negative for G. destructans but positive for other fungi...
After direct PCR amplification and DNA sequence analysis of fungal rRNA gene ITS regions, genetic signatures 100% identical with those from G. destructans type isolate NWHC 20631–21 (GenBank accession no. EU884921) were identified from 21 of 23 bats examined: 15/15 from Germany, 2/2 from Hungary, and 4/4 from Switzerland. Both bats from the United Kingdom were colonized by Penicillium sp. (Tables 1, 2). Fungi with conidia morphologically identical to those of G. destructans (Figure 1, panel B) as described by Gargas et al. (6) were isolated in axenic cultures from 8 of 23 bats examined: 3/15 from Germany, 1/2 from Hungary, and 4/4 from Switzerland) (Tables 1, 2; Figure 2).
Consistent with published descriptions for G. destructans (6), fungal colonies grew slowly and within 14 days attained diameters of 1.0 mm at 4°C and 4.0–5.0 mm at 8°C; no growth occurred at 25°C. The sensitivity of our method for isolating G. destructans from bat hair was comparable to published diagnostic sensitivity for culturing G. destructans from bat skin (20). Subsequent PCR/DNA sequencing analyses of the 8 isolates indicated that they all had rRNA gene ITS and SSU region DNA sequences identical to those of G. destructans type isolate NWHC 20631–21 (GenBank accession nos. EU884921 for ITS and FJ231098 for SSU).
Unlike other bats sampled in this study, the 2 greater horseshoe bats from the United Kingdom were found dead, and their nostrils were colonized by Penicillium sp. These bats did not fulfill the pathologic criteria for WNS (7) because fungal hyphae did not invade the epidermis but remained within the superficial layer of the epidermal stratum corneum. A more complete description of the postmortem analysis of the greater horseshoe bats has been reported (21). G. destructans was not isolated in culture, and its genetic signature was not identified by PCR and DNA sequencing of samples collected from greater horseshoe bats.
Laboratory analyses demonstrated that 5 species of the genus Myotis in Europe harbored G. destructans; male and female bats were equally affected. Despite laboratory confirmation that bats obtained in Germany, Switzerland, and Hungary were colonized by G. destructans, deaths were not observed at collection sites. Puechmaille et al. (16) reported a similar observation with a greater mouse-eared bat in France. Additionally, a lesser mouse-eared bat from Hungary with visible fungal infection during hibernation, from which G. destructans was isolated, was recaptured 5 months later (August 2009) and showed no external signs of fungal infection. On February 19, 2010, the same bat was again observed in the same hibernaculum without any visible sign of fungal growth. However, 7 other bats within that group of 55 animals displayed obvious fungal growth but were not sampled for this study.
In contrast, decreases in bat hibernating colonies infected by G. destructans in North America are often >90% (2,3), and mortality rates similar in magnitude would be difficult to miss among closely monitored winter populations of bats in Europe. Biologists in Germany and Switzerland have conducted annual censuses of bat hibernacula since the 1930s and 1950s, respectively. In Hungary, the largest hibernacula have been annually monitored since 1990. Similar death rates to those caused by WNS in hibernating bats in North America have never been documented in countries in Europe in which G. destructans has now been identified.
Although distribution of G. destructans in bats across Europe has not been exhaustively characterized, opportunistic sampling conducted as part of this study during the winter of 2008–09 demonstrated that the fungus was present on bats in 3 countries (Figure 2). The 2 most distant points from which bats colonized with G. destructans have been identified were separated by >1,300 km. Despite the observed distribution of G. destructans in Europe (Figure 2), the 5 bat species from which G. destructans was detected migrate average distances <100 km between their summer and winter roosting sites (22), indicating that the fungus is most likely spread as local bat populations emerge from hibernacula, disperse, and interact with populations within their dispersal range. Identification of bats colonized by G. destructans from such distant sites, in addition to the relatively homogenous distribution of the fungus among sites in Germany, suggests that G. destructans may be widespread in Europe.
Regardless of widespread occurrence of G. destructans among bat species in Europe (Figure 2), deaths of bats in Europe caused by WNS, similar to those caused by WNS in North America, have not been observed. Although no bat species migrates between Europe and North America or is present on both continents (23,24), many species of the genus Myotis are infected by G. destructans on each continent. Although the mechanism(s) by which hibernating bats died because of infection with G. destructans in North America is not yet understood, bat species in Europe may exhibit greater resistance or respond differently to infection by this fungus than their counterparts in North America.
Before the emergence of WNS in North America, large aggregations of hibernating bats ranging from 1,000 to 50,000 animals were common in caves and mines of affected regions, and many hibernation sites in regions of North America still unaffected by WNS contained tens of thousands of bats during winter (some contain hundreds of thousands) (25). In contrast, aggregations of bats hibernating in caves and mines in Europe rarely exceed 1,000 animals. However, larger hibernating groups have been observed at a few natural sites, such as a cave in northern Germany with 13,000–18,000 bats (26) and human-made structures (e.g., Daubenton bats in bunkers and catacombs) (24). If host density plays a role in G. destructans transmission or deaths of bats, such as through increased disturbance of clustered bats, the bats in Europe may experience lower mortality rates because they form smaller hibernation groups composed of small clusters or individual bats. Apparent continental differences in susceptibility of hibernating bats to deaths associated with skin infection by G. destructans may indicate either circumstantial or evolved resistance in bats in Europe.
G. destructans has been detected in North America only in states and provinces where WNS has also been observed and in contiguous states. Recent emergence and spread of G. destructans with associated deaths of bats throughout hibernacula in the northeastern United States (3) may suggest ecologic release of an exotic pathogen into an uninfected ecosystem. Although this suggestion remains a hypothesis and how G. destructans may have been introduced to the United States is not known, initial documentation of WNS in a popular tourist cave near Albany, New York (1), suggests that a human vector could have been involved.
There are many examples of unintended introductions of fungal pathogens, particularly of those affecting plants and ectothermic animals with tissue temperatures permissive to fungal infection (27–29). One case with striking similarities is the panzootic chytrid fungus (Batrachochytrium dendrobatidis), which has caused global decreases among amphibian species (30). As with skin infection by B. dendrobatidis in amphibians, which can alter body electrolyte levels and lead to cardiac arrest (31), skin infection by G. destructans in hibernating bats may also kill by causing irreversible homeostatic imbalance because wing membranes play major roles in water balance, circulation, and thermoregulation of hibernating bats during winter (32,33).
Bat species in Europe may be immunologically or behaviorally resistant to G. destructans because of having coevolved with the fungus. Additionally, microbial flora of bat skin or other abiotic surfaces in bat hibernacula in Europe may have also coevolved to incorporate G. destructans as a nonpathogenic component of the microbial community. Conversely, possible recent introduction of G. destructans into the United States, with subsequent infection of bat species in North America and ecosystems not infected with the fungus, provides a potential explanation for the devastating effects of WNS in North America. Although bats are reservoirs of various pathogens (34,35), research into the immune function of bats, particularly during hibernation, is just beginning.
In conclusion, nondetrimental colonization of bat species in Europe by G. destructans may be relatively common (Figure 2), and historical reports (15) suggest that such colonization of hibernating bats in Europe has occurred for several decades. In contrast to recent mass deaths associated with G. destructans skin infection, which is the hallmark of WNS in North America, bats in Europe appear to coexist with G. destructans. Studies to investigate mechanisms of pathogenesis, microbial ecology, and phylogeography of G. destructans will be essential for developing a comprehensive understanding of WNS. In particular, testing the hypotheses that bats in Europe are more resistant to fungal skin infection by G. destructans, that G. destructans was introduced from Europe to North America, and that environmental circumstances limit the pathogenicity of G. destructans in Europe seem warranted. Divergent manifestations of infection by G. destructans in bats in Europe and North America provide a unique opportunity to address these research objectives with the ultimate goals of better understanding WNS and developing sound strategies to manage the devastating effects of this emerging wildlife disease in North America.
Dr Wibbelt is a senior veterinary pathologist at the Leibniz Institute for Zoo and Wildlife Research, Berlin, Germany. Her research interests include infectious diseases in wild animals, particularly bats.
Acknowledgments
We thank N. Jahn, D. Viertel, A. Kus, and C. Kohl for excellent technical assistance; A. Beck, T. Filip, M. Franz, S. Gloor, R. Güttinger, A. Kiefer, V. Korn, G. Mäscher, B. Máté, C.-D. Morawitz, C. Morris, P. Paulovics, M. Piskorski, F. Putzmann, W. Schorcht, and C. Tress for help surveying sites and retrieving samples; A. Gargas, K. Schuler, and 2 anonymous reviewers for providing thoughtful comments on earlier drafts of the manuscript; and M. Riccucci for help with the literature search for previous reports of fungi in bats in Europe.
This study was supported by Bat Conservation Switzerland.
References
- Blehert DS, Hicks AC, Behr M, Meteyer CU, Berlowski-Zier BM, Buckles EL, Bat white-nose syndrome: an emerging fungal pathogen? Science. 2009;323:227. DOIPubMedGoogle Scholar
- Reichard JD, Kunz TH. White-nose syndrome inflicts lasting injuries to the wings of little brown myotis (Myotis lucifugus). Acta Chiropt. 2009;11:457–64. DOIGoogle Scholar
- Turner GR, Reeder DM. Update of white-nose syndrome in bats, September 2009. Bat Research News. 2009;50:47–53.
- Courtin F, Stone W, Risatti G, Gilbert K, Van Kruiningen H. Pathologic findings and liver elements in hibernating bats with white-nose syndrome. Vet Pathol. 2010;47:214–9. DOIPubMedGoogle Scholar
- United States Geological Survey. Update on white-nose syndrome: Tennessee finding. USGS wildlife health bulletin. Reston (VA): The Survey; 2010 [cited 2010 May 19]. http://www.nwhc.usgs.gov/disease_information/white-nose_syndrome/
- Gargas A, Trest MT, Christensen M, Volk TJ, Blehert DS. Geomyces destructans sp. nov. associated with bat white-nose syndrome. Mycotaxon. 2009;108:147–54. DOIGoogle Scholar
- Meteyer CU, Buckles EL, Blehert DS, Hicks AC, Green DE, Shearn-Bochsler V, Histopathologic criteria to confirm white-nose syndrome in bats. J Vet Diagn Invest. 2009;21:411–4.PubMedGoogle Scholar
- Chabasse D, Guiguen C, Couatarmanac’h A, Launay H, Reecht V, de Bièvre C. Keratinophilic fungal flora isolated from small wild mammals and rabbit-warren in France. Discussion on the fungal species found [in French]. Ann Parasitol Hum Comp. 1987;62:357–68.PubMedGoogle Scholar
- Mercantini R, Marsella R, Prignano G, Moretto D, Marmo W, Leonetto F, Isolation of keratinophilic fungi from the dust of ferry boats and trains in Italy. Mycoses. 1989;32:590–4. DOIPubMedGoogle Scholar
- de Bellis T, Kernaghan G, Widden P. Plant community influences on soil microfungal assemblages in boreal mixed-wood forests. Mycologia. 2007;99:356–67. DOIPubMedGoogle Scholar
- Kochkina GA, Ivanushkina NE, Akimov VN, Gilichinskii DA, Ozerskaia SM. Halo- and psychrotolerant Geomyces fungi from arctic cryopegs and marine deposits [in Russian]. Mikrobiologiia. 2007;76:39–44.PubMedGoogle Scholar
- Mercantini R, Marsella R, Cervellati M. Keratinophilic fungi isolated from Antarctic soil. Mycopathologia. 1989;106:47–52. DOIPubMedGoogle Scholar
- Gianni C, Caretta G, Romano C. Skin infection due to Geomyces pannorum var. pannorum. Mycoses. 2003;46:430–2. DOIPubMedGoogle Scholar
- Zelenková H. Geomyces pannorum as a possible causative agent of dermatomycosis and onychomycosis in two patients. Acta Dermatovenerol Croat. 2006;14:21–5.PubMedGoogle Scholar
- Feldmann R. Teichfledermaus–Myotis dasycneme (Boie, 1825). Die Säugetiere Westfalens. Münster: Westfälisches Museum für Naturkunde; 1984. p. 107–11.
- Puechmaille SJ, Verdeyroux P, Fuller H, Ar Gouilh M, Bekaert M, Teeling EC. White-nose syndrome fungus (Geomyces destructans) in bat, France. Emerg Infect Dis. 2010;16:290–3.PubMedGoogle Scholar
- White T, Bruns T, Lee S, Taylor J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TH, editors. PCR protocols: a guide to methods and applications. San Diego (CA): Academic Press; 1990. p. 315–22.
- Gargas A, Taylor J. Polymerase chain reaction (PCR) primers for amplifying and sequencing 18S rDNA from lichenized fungi. Mycologia. 1992;84:589–92. DOIGoogle Scholar
- Gargas A, dePriest P, Taylor J. Positions of multiple insertions in SSU rDNA of lichen forming fungi. Mol Biol Evol. 1995;12:208–18.PubMedGoogle Scholar
- Lorch JM, Gargas A, Meteyer CU, Berlowski-Zier BM, Green DE, Shearn-Bochsler V, Rapid polymerase chain reaction diagnosis of white-nose syndrome in bats. J Vet Diagn Invest. 2010;22:224–30.PubMedGoogle Scholar
- Barlow A, Ford S, Green R, Morris C, Reaney S. Investigations into suspected white-nose syndrome in two bats in Somerset. Vet Rec. 2009;165:481–2. DOIPubMedGoogle Scholar
- Hutterer R, Ivanova T, Meyer-Cords C, Rodrigues L. Bat migrations in Europe: a review of banding data and literature. Bonn (Germany): German Agency for Nature Conservation; 2005.
- Nowak R. Walker’s mammals of the world. Baltimore: The Johns Hopkins University Press; 1999.
- Dietz C, von Helversen O, Nill D. Bats of Britain, Europe and Northwest Africa. London: A and C Black Publishers; 2009.
- Barbour RW, Davis WH. Bats of America. Lexington (KY): The University Press of Kentucky; 1969.
- Petermann R, Boye P. National report on bat conservation in the Federal Republic of Germany 2003–2006. Bonn (Germany): Eurobats; 2006.
- Casadevall A. Fungal virulence, vertebrate endothermy, and dinosaur extinction: is there a connection? Fungal Genet Biol. 2005;42:98–106. DOIPubMedGoogle Scholar
- Desprez-Loustau ML, Robin C, Buée M, Courtecuisse R, Garbaye J, Suffert F, The fungal dimension of biological invasions. Trends Ecol Evol. 2007;22:472–80. DOIPubMedGoogle Scholar
- Robert VA, Casadevall A. Vertebrate endothermy restricts most fungi as potential pathogens. J Infect Dis. 2009;200:1623–6. DOIPubMedGoogle Scholar
- Fisher MC, Garner TW, Walker SF. Global emergence of Batrachochytrium dendrobatidis and amphibian chytridiomycosis in space, time, and host. Annu Rev Microbiol. 2009;63:291–310. DOIPubMedGoogle Scholar
- Voyles J, Young S, Berger L, Campbell C, Voyles WF, Dinudom A, Pathogenesis of chytridiomycosis, a cause of catastrophic amphibian declines. Science. 2009;326:582–5. DOIPubMedGoogle Scholar
- Davis WH. Hibernation: ecology and physiological ecology. In: Wimsatt WA, editor. Biology of bats. Vol. 1. New York: Academic Press; 1970. p. 265–300.
- Makanya AN, Mortola JP. The structural design of the bat wing web and its possible role in gas exchange. J Anat. 2007;211:687–97. DOIPubMedGoogle Scholar
- Calisher CH, Childs JE, Field HE, Holmes KV, Schountz T. Bats: an important reservoir host of emerging viruses. Clin Microbiol Rev. 2006;19:531–45. DOIPubMedGoogle Scholar
- Wibbelt G, Speck S, Field H. Methods for assessing diseases in bats. In: Kunz T, Parsons S, editors. Ecological and behavioral methods for the study of bats. Baltimore: Johns Hopkins University Press; 2009. p. 775–94.
Figures
Tables
Cite This ArticleTable of Contents – Volume 16, Number 8—August 2010
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Gudrun Wibbelt, Wildlife Diseases Pathology Group, Leibniz Institute for Zoo and Wildlife Research, Alfred-Kowalke-Strasse 17, 10315 Berlin, Germany
Top