Volume 19, Number 11—November 2013
Research
CTX-M β-Lactamase–producing Klebsiella pneumoniae in Suburban New York City, New York, USA
Cite This Article
Citation for Media
Abstract
CTX-M extended-spectrum β-lactamase (ESBL)–producing Klebsiella pneumoniae isolates are infrequently reported in the United States. In this study, we analyzed nonduplicate ESBL-producing K. pneumoniae and Escherichia coli clinical isolates collected during 2005–2012 at a tertiary care medical center in suburban New York City, USA, for the presence of blaCTX-M, blaSHV, blaTEM, and blaKPC genes. Despite a high prevalence of blaCTX-M genes in ESBL-producing E. coli since 2005, blaCTX-M genes were not detected in K. pneumoniae until 2009. The prevalence of CTX-M–producing K. pneumoniae increased significantly over time from 1.7% during 2005–2009 to 26.4% during 2010–2012 (p<0.0001). CTX-M-15 was the dominant CTX-M genotype. Pulsed-field gel electrophoresis and multilocus sequence typing revealed high genetic heterogeneities in CTX-M–producing K. pneumoniae isolates. This study demonstrates the recent emergence and polyclonal spread of multidrug resistant CTX-M–producing K. pneumoniae isolates among patients in a hospital setting in the United States.
CTX-M enzymes are a group of class A extended-spectrum β-lactamases (ESBLs) that are rapidly spreading among Enterobacteriaceae worldwide (1). Since the initial isolation of CTX-M-1 from a European patient in the late 1980s (2), >130 CTX-M allelic variants have been described (http://www.lahey.org/Studies/other.asp#table1). These CTX-M variants have been divided into 5 major phylogenetic groups, CTX-M-1, CTX-M-2, CTX-M-8, CTX-M-9, or CTX-M-25 on the basis of their amino acid sequences (1,2).
During the past decade, CTX-M enzymes have become the most prevalent ESBL enzymes in clinical Enterobacteriaceae isolates, especially in ESBL-producing Escherichia coli in Europe, Asia, and South America (1,3). By contrast, SHV- and TEM-type ESBL enzymes are primarily found in ESBL-producing K. pneumoniae and E. coli clinical isolates in North America (3). In the United States, CTX-M–like ESBL-producing Enterobacteriaceae was first reported in 2003, when CTX-M enzymes were detected in 9 E. coli clinical isolates from 5 US states (4). The spread of CTX-M type ESBL in Enterobacteriaceae, however, was not appreciated until 2007 when a Texas study showed a high prevalence of CTX-M ESBL in E. coli clinical isolates recovered during 2000–2005 (5). Since then, CTX-M–producing E. coli isolates have been documented in dispersed US geographic regions (3,6,7). CTX-M enzymes are now the predominant ESBL type in E. coli clinical isolates in Texas (5), Pennsylvania (6), Illinois (8), and New York (9,10).
CTX-M–type ESBL enzymes have also been reported in the United States in some non–E. coli Enterobacteriaceae species, such as Klebsiella spp. (5,11,12), Proteus mirabilis (5,11), Enterobacter spp. (5), Salmonella spp. (13), Shigella spp. (14), and Morganella morganii (5). Nevertheless, CTX-M–type ESBL have been principally detected and reported in E. coli clinical isolates. To date, <50 CTX-M–producing K. pneumoniae isolates have been described in the United States, and the epidemiologic and microbiological data provided have been limited (5,11,12,15–18). The implications of CTX-M–producing K. pneumoniae for laboratory detection, patient care, and public health in the United States remain to be elucidated.
In this study, we investigated the prevalence of SHV-, TEM-, and CTX-M–encoding genes in a large collection of ESBL-producing K. pneumoniae and E. coli clinical isolates from a tertiary care medical center in suburban New York City in Westchester County, New York, over an 8-year period (2005–2012). Microbiological characteristics of CTX-M ESBL-producing K. pneumoniae isolates were examined, and certain clinical/epidemiologic features of patients with these isolates were analyzed.
Bacterial Isolates and Phenotypic Detection of ESBLs
Nonduplicate K. pneumoniae clinical isolates were recovered from patient specimens during January 2005–July 2012 at the clinical microbiology laboratory of Westchester Medical Center. These included 208 blaKPC-negative non-K. pneumoniae carbapenemase (non-KPC) ESBL-producing or third-generation cephalosporin-resistant K. pneumoniae isolates and 228 KPC (blaKPC-positive)–producing K. pneumoniae isolates. In addition, 163 nonduplicate ESBL-producing E. coli clinical isolates from the same period were also analyzed for comparison. Isolates were randomly selected to span the entire study year with an approximately equal number of isolates per quarter; only 1 isolate from each patient was chosen and tested. The center is a 643-bed academic tertiary-care medical center in Westchester County, New York. The Institutional Review Board of New York Medical College approved this study.
The bacterial isolates were identified and evaluated for antimicrobial drug susceptibility with the MicroScan WalkAway 96 system (Siemens, Sacramento, CA, USA). ESBL production was phenotypically confirmed by a double-disk or broth microdilution method for suspected ESBL isolates according to the Clinical and Laboratory Standards Institute (CLSI) guidelines (19). The antimicrobial drug susceptibility of CTX-M–producing K. pneumoniae isolates against selected antimicrobial drugs was also assessed with standardized CLSI disk diffusion and Etest methods. Bacterial isolates were refrigerated on nutrient agar slants or were frozen (−80°C) in MicroBank cryovials containing 20% glycerol (Pro-Lab Diagnostics, Round Rock, TX, USA). For antimicrobial drug susceptibility testing of frozen isolates, fresh subcultures were used per CLSI guidelines.
PCR Detection of blaCTX-M, blaSHV, blaTEM, and blaKPC Genes
For PCR, bacterial genomic DNA was extracted directly from colonies on nutrient slants or from fresh subcultures grown on Trypticase soy agar with 5% sheep blood (TSA II, BBL, Sparks, MD, USA) by boiling a dense suspension of an approximately no. 1 McFarland standard in sterile distilled water. As the DNA template in the PCR assays, 2–3 μL of the boiled cell suspension was used. PCR amplification of blaCTX-M, blaSHV, blaTEM, and blaKPC genes in K. pneumoniae and E. coli clinical isolates was performed by using a consensus primer pair specific to each type of β-lactamase as described (20–22). A multiplex PCR was developed and used for simultaneous detection of blaCTX-M (551 bp) and blaTEM (972 bp) genes. Two PCRs were performed for blaSHV-ESBL and blaKPC, respectively. PCRs were carried out by using the HotStart DNA polymerase master mix (QIAGEN, Germantown, MD, USA) with 30–35 cycles at an annealing temperature of 50°C for blaCTX-M and blaTEM, and 52°C for blaSHV and blaKPC. PCR products were analyzed by agarose gel electrophoresis or by using the QIAxcel system (QIAGEN). The specificity of PCR amplicons on representative isolates was confirmed by DNA sequencing.
DNA Sequencing
For DNA sequencing, PCR products were purified by using the PCR Purification kit (QIAGEN) or the ExoSAP-IT PCR Clean-up kit (Affymetrix, Cleveland, OH, USA), according to the manufacturer’s instructions. The purified DNA amplicons were sequenced by using an ABI Prism BigDye Terminator (version 1.1) cycle sequencing ready reaction kit on the ABI Prism 3730xl or ABI 3500xl DNA Analyzers (Applied Biosystems, Foster City, CA, USA) in-house, or by a commercial facility (GeneWiz, South Plainfield, NJ, USA). The CTX-M, TEM, and SHV gene sequences were compared with sequences in GenBank by using the NCBI basic local alignment search tool (www.ncbi.nlm.nih.gov/BLAST).
Multilocus Sequence Typing
Multilocus sequence typing (MLST) was performed by using primers and conditions as described by Diancourt et al. (23). PCR products from MLST were sequenced as described above. Allelic profiling and sequence types (STs) were determined by querying the K. pneumoniae MLST database maintained by the Pasteur Institute (www.pasteur.fr/recherche/genopole/PF8/mlst/Kpneumoniae.html).
Pulsed-field Gel Electrophoresis (PFGE)
Pulsed-field gel electrophoresis (PFGE) on CTX-M ESBL–producing K. pneumoniae isolates representing each CTX-M genotype was performed as described (24). The GelCompare II software, (version 2.0; Applied Maths, Austin, TX, USA) was used to calculate the Dice similarity coefficients and generate dendrograms by cluster analysis with the unweighted-pair group method using average linkages. Pulsotype designations were assigned at the ≥80% profile similarity level.
CTX-M in ESBL-producing, non-KPC K. pneumoniae Clinical Isolates
Of the 121 ESBL-producing K. pneumoniae isolates originally recovered during 2005–2009, blaSHV and blaTEM genes were detected in 102 (84.3%) and 61 (50.4%) of 121 isolates, respectively (Table 1). Overall, 25 CTX-M-type ESBL K. pneumoniae were identified. However, none of the 81 K. pneumoniae isolates from 2005 through 2008 was positive for blaCTX-M genes. CTX-M–type ESBL was first detected in 2 (5.0%) of 40 K. pneumoniae isolates from 2009. The prevalence of K. pneumoniae isolates carrying the CTX-M–encoding genes increased to 6 (17.6%) of 34 in 2010 and 12 (34.3%) of 35 in 2011. The level remained high (27.8%, 5/18) in the first 7 months of 2012. Overall, only 2 (1.7%) of 121 ESBL-producing K. pneumoniae isolates from 2005 through 2009 carried the blaCTX-M genes, compared with 23 (26.4%) of 87 isolates from 2010 through 2012 (p<0.0001, Fisher exact test), indicating the rapid emergence and spread of CTX-M enzymes among ESBL-producing K. pneumoniae clinical isolates since 2009.
CTX-M in ESBL-producing E. coli Clinical Isolates
One hundred sixty-three ESBL-producing E. coli clinical isolates from 2005 through 2012 were analyzed by PCR for detection of blaESBL genes of the SHV, TEM, and CTX-M types (Table 2). Unlike the situation with K. pneumoniae, blaCTX-M genes were detected in ESBL-producing E. coli isolated as early as 2005. Overall, 89 (54.6%) of 163 ESBL E. coli isolates from the 8-year period carried blaCTX-M genes. CTX-M was the leading ESBL type in all years examined except 2008. The blaCTX-M genes from 47 (52.8%) of 89 CTX-M–producing E. coli isolates were sequenced. CTX-M-15 was determined in 45 (95.7%) of 47 CTX-M–producing E. coli isolates analyzed. CTX-M-1 and CTX-M-3 genotypes were each found in 1 ESBL E. coli isolate.
CTX-M in KPC-producing K. pneumoniae Clinical Isolates
Two hundred twenty-eight KPC-producing K. pneumoniae isolates from 2005 to 2012 were examined by PCR for detection of blaCTX-M genes. All K. pneumoniae isolates were positive for the blaKPC gene by PCR as described (22). None was positive for the blaCTX-M gene.
Clinical and Microbiological Characteristics of CTX-M–producing K. pneumoniae
Selected clinical/epidemiologic features of the 25 patients with CTX-M ESBL–producing K. pneumonia and certain microbiological characteristics of the isolates are shown in Table 3, Appendix. Mean patient age was 56 years, and 13 (52%) of the patients were male. Sixteen patients (64%) had bacteriuria. CTX-M–producing K. pneumoniae isolates were recovered from 13 (52%) patients within 72 hours of hospital admission; however, 18 (72%) of these patients had been hospitalized in the 8 months before the current admission.
The blaCTX-M genes from all 25 CTX-M ESBL–producing K. pneumoniae isolates from 2009 through 2012 were sequenced. CTX-M-15 was identified in 19 (76.0%) and was the dominant CTX-M genotype. The remaining 6 isolates were determined to be CTX-M-3 (n = 4), CTX-M-1 (n = 1), and CTX-M-2 (n = 1), respectively. Twenty-four (96.0%) had coexisting blaSHV β-lactamases, which were predominantly non-ESBL blaSHV-11 (n = 15) and blaSHV-1 (n = 5). Four additional K. pneumoniae carried ESBL-type blaSHV β-lactamases, including blaSHV-12 (n = 1), blaSHV-27 (n = 1), and blaSHV-28 (n = 2). Seventeen (68.0%) were positive for TEM-type β-lactamases, and all were confirmed to be blaTEM-1.
The antimicrobial drug susceptibilities of CTX-M–producing K. pneumoniae isolates are summarized in Table 4. Of the 25 CTX-M–producing K. pneumoniae isolates examined in this study, only 12% (n = 3) and 36% (n = 8) of isolates were susceptible to ciprofloxacin and gentamicin, respectively. Low susceptibility rates were also observed for pipercillin/tazobactam (36%), tetracycline (20%) and trimethoprim/sulfamethoxazole (4%). Twenty-three of the 25 (92%) isolates tested were susceptible to carbapenems. Notably, the 2 carbapenem-resistant K. pneumoniae isolates (PK30 and PK107) carried blaCTX-M-3, blaSHV-11, and blaTEM-1. One of these K. pneumoniae isolates also showed resistance to colistin with an MIC of 64µg/mL. Both patients died of complications associated with bloodstream and respiratory tract infections. Three of 22 CTX-M–producing K. pneumoniae isolates examined by Etest were nonsusceptible to tigecycline (MICs 3 µg/mL, 3 µg/mL, and 8 µg/mL).
Figure 1
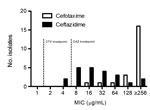
Figure 1. . MIC distribution for cefotaxime (CTX) and ceftazidime (CAZ) in CTX-M extended-spectrum β-lactamase–producing Klebsiella pneumoniae clinical isolates from a tertiary care medical center in suburban New York, NY, USA, 2005–2012 (n...
All 25 CTX-M–producing K. pneumoniae isolates examined were resistant to cefotaxime, and all but 1 isolate showed higher MICs of cefotaxime than of ceftazidime. The 50% minimum inhibitory concentration (MIC50) for cefotaxime among these isolates was >256 µg/mL. By contrast, the MIC50 and 90% inhibitory concentration for ceftazidime were 16 µg/mL and 128 µg/mL, respectively. Two CTX-M–producing K. pneumoniae isolates (8.0%) were susceptible (MIC ≤4 µg/mL) and 5 isolates (20%) were intermediate in susceptibility (8 µg/mL) to ceftazidime according to the 2010 revised CLSI breakpoints (Figure 1). In addition, we determined the susceptibilities of 22 CTX-M–producing K. pneumoniae isolates against cefotaxime and ceftazidime by using the standard disk diffusion method. All CTX-M–producing K. pneumoniae isolates examined were resistant to cefotaxime by disk diffusion (mean inhibitory zone size 8.3 mm; range 6–13 mm). Two of these isolates were susceptible (≥21 mm) and 5 had intermediate (18–20 mm) susceptibility to ceftazidime by disk diffusion (Table 3, appendix). The disk diffusion results showed a category agreement with the Etest MIC of 100% for cefotaxime and 90.9% for ceftazidime with 2 minor errors.
PFGE and MLST Analysis of CTX-M–producing K. pneumoniae
Figure 2

Figure 2. . Dendrogram of pulsed-field gel electrophoresis (PFGE) patterns showing the genetic relatedness of CTX-M extended-spectrum β-lactamase (ESBL)–producing Klebsiella pneumoniae isolates from patients in suburban New York, NY, USA (n = 17)....
Of 17 representative CTX-M-producing K. pneumoniae isolates analyzed by PFGE, 8 different pulsotypes (PF1–8) were identified with Dice coefficients of ≥80% similarity (Figure 2). Ten of 17 K. pneumoniae isolates examined belonged to 3 major groups (PF3, PF4, PF5) with 3–4 isolates in each group. The remaining pulsotypes contained only 1 or 2 K. pneumoniae isolates. No clear temporal relationship was shown among the highly related isolates.
MLST was performed on 18 CTX-M–producing K. pneumoniae isolates. These isolates were selected to represent different CTX-M genotypes, pulsotypes, antimicrobial susceptibility profiles, and years of isolation. Twelve STs were recognized for the K. pneumoniae isolates examined (Table 3, appendix). Notably, all 3 CTX-M group 1, non–CTX-M-15 K. pneumoniae isolates analyzed (KP38, PK107, and PK135) had ST11, whereas 10 different STs (ST15, ST16, ST17, ST48, ST147, ST252, ST258, ST280, ST392, and ST437) were identified for the 14 CTX-M-15 K. pneumoniae isolates. Isolate F351 was the only non–CTX-M-1 group K. pneumoniae isolate identified in this study and was determined to be a separate group (ST792) by MLST. Of the 14 CTX-M–producing K. pneumoniae isolates evaluated simultaneously by DNA sequencing, PFGE and MLST, a high genetic divergence was demonstrated by the detection of 4 CTX-M genotypes (CTX-M-1, CTX-M-2, CTX-M-3, and CTX-M-15), 8 pulsotypes (PF1–8) and 11 STs (ST11, ST15, ST16, ST17, ST48, ST147, ST252, ST280, ST392, ST437, and ST792) (Figure 2).
CTX-M ESBL–producing E. coli, especially ST131 strains, have emerged in recent years in several US states (5–7,25,26). In this study, we detected blaCTX-M genes in ESBL-producing E. coli strains isolated from patients at a tertiary care medical center in suburban New York City as early as 2005. Eighty-nine (54.6%) of 163 ESBL-producing E. coli isolates in the study period (2005–2012) carried blaCTX-M. Our findings confirm the emergence and dominance of CTX-M enzymes in ESBL-producing E. coli since the mid-2000s in the New York City metropolitan area (9,10).
Despite this high prevalence of CTX-M in ESBL-producing E. coli since 2005, none of 81 ESBL-producing K. pneumoniae isolates recovered from patients at the same tertiary care medical center from 2005 through 2008 was positive for blaCTX-M. CTX-M-type ESBL was first detected in K. pneumoniae isolates from our institution in 2009. The percentage of K. pneumoniae isolates carrying blaCTX-M has increased significantly since then. During 2010–2012, blaCTX-M genes were identified in 23 of 87 (26.4%) ESBL-producing K. pneumoniae isolates. These data demonstrate the rapid emergence and spread of CTX-M ESBL-producing K. pneumoniae in our patients. To date, CTX-M–producing K. pneumoniae has been recognized in several US states, including Texas (2004–2007, n = 11) (5,12), Nebraska (2005, n = 1) (15), Pennsylvania (2007, n = 5) (11), and 1 isolate in 2007 each from California, Massachusetts, Michigan, New Jersey, New York, Washington, and Wisconsin (12). In addition, a few CTX-M K. pneumoniae isolates have been reported from 2 collections of the SMART surveillance program with isolates recovered during 2008–2009 (16) and 2009–2010 (18). No CTX-M was detected in US ESBL-producing K. pneumoniae isolates collected before 2000 (3), with all CTX-M–producing K. pneumoniae isolates recovered from US patients in or after 2004. Therefore, we speculate that the emergence and spread of blaCTX-M in K. pneumoniae are recent evolutionary events that most likely occurred in the mid- to late-2000s in the United States.
The particular CTX-M enzyme type in ESBL-producing K. pneumoniae and E. coli varies geographically. CTX-M-15, which belongs to the CTX-M-1 group, is the most prevalent CTX-M allele with a worldwide distribution (1,2,26). CTX-M-14, which belongs to the CTX-M-9 group, is another common variant that is highly prevalent in some European and Asian countries (27–30), whereas CTX-M-2 in the CTX-M-2 group and CTX-M-8 seem to be dominant in South America (1,31). In the United States, CTX-M-15 is the most frequently detected genotype among CTX-M–producing K. pneumoniae isolates, followed by CTX-M-14 (5,11,12). CTX-M-2 group and CTX-M-8 group ESBL-producing K. pneumoniae each was identified in 1 isolate (16).
Our data provide strong evidence for the recent, rapid emergence, and polyclonal spread of the CTX-M-1 group, especially CTX-M-15 ESBL-producing K. pneumoniae in a US hospital setting. In this study, 24 (96.0%) of 25 blaCTX-M-positive K. pneumoniae were categorized as group 1 CTX-M, including isolates encoding CTX-M-15 (n = 19), CTX-M-1 (n = 1), and CTX-M-3 (n = 4). Similarly, group 1 CTX-M was detected in 47 (100%) of 47 blaCTX-M-positive E. coli isolates. In addition, 1 K. pneumonia isolate had the CTX-M-2 genotype. No CTX-M-14 was detected in these K. pneumoniae and E. coli isolates. CTX-M-14 has been reported in E. coli ESBL isolates in several US states, including geographically adjacent Pennsylvania (6). CTX-M-14 has also been reported in K. pneumoniae isolates in the Calgary Healthcare Region of Canada (32). Why CTX-M-14 is absent in the ESBL-producing E. coli and K. pneumoniae isolates from the New York, NY metropolitan area is unknown. Because CTX-M-15–producing K. pneumoniae isolates may exhibit significantly higher resistance rates to ciprofloxacin and pipercillin-tazobactam than CTX-M-14–producing isolates (27,28), CTX-M genotypes and their antimicrobial drug profiles should be monitored among CTX-M–producing E. coli and K. pneumoniae isolates in regions where they are emerging.
We investigated the genetic relatedness of CTX-M–producing K. pneumoniae isolates by PFGE and MLST. Of the 17 representative isolates examined by PFGE, 8 different pulsotypes were determined. Similarly, 12 MLST STs were identified for the 18 CTX-M–producing isolates analyzed. Our data, in combination with findings from other groups (1), suggest that CTX-M–producing K. pneumoniae isolates are genetically heterogeneous. The emergence and polyclonal spread of CTX-M–producing K. pneumoniae likely occurred among isolates with diverse genetic backgrounds. This hypothesis contrasts with findings regarding KPC-producing K. pneumoniae: a clonal spread of KPC-producing K. pneumoniae isolates belonging to the ST258 lineage was observed by us (33) and Pournaras et al. (34). In clinical strains, CTX-M–encoding genes have commonly been located on plasmids that vary in size from 7 kb to 160 kb (2). Plasmid-mediated transmission of CTX-M genes in Enterobacteriaceae that involves several motile genetic elements has been described (2,35,36). Given the dominance of CTX-M-15 genotypes among genetically heterogeneous K. pneumoniae isolates, our study also implies the probable horizontal transfer of a genetic element carrying blaCTX-M among K. pneumoniae isolates.
Of the 12 STs determined for the CTX-M ESBL–producing K. pneumoniae isolates, ST11, ST15, ST17, ST48, ST147, and ST258 have been reported in CTX-M–positive K. pneumoniae in Spain, Hungary, or Korea (28,37,38). Among these, only ST17 was reported among CTX-M–producing K. pneumoniae isolates in Canada (39). In this study, we determined the STs among CTX-M–producing K. pneumoniae in the United States and document the existence of 6 STs (ST16, ST252, ST280, ST392, ST437, ST792) in CTX-M–producing K. pneumoniae not previously described.
The CTX-M–producing K. pneumoniae isolates evaluated in this study showed several notable epidemiologic, clinical, and microbiological features. First, most CTX-M–producing isolates were recovered from patients with bacteriuria, which is similar to that observed for infections caused by CTX-M–producing E. coli in New York City (9,10). Although CTX-M–producing K. pneumoniae was isolated in clinical specimens collected within 72 hours of hospitalization in about half of the patients, 18 (72%) of 25 patients had been hospitalized in the prior 8 months. This factor highlights the potential for acquiring CTX-M–producing K. pneumoniae in health care settings and differs from the experience with CTX-M–producing E. coli that are associated with infections arising in the community setting unrelated to exposure to health care facilities (26). Second, the CTX-M–producing K. pneumoniae study isolates exhibited high rates of resistance to gentamicin (68%), trimethoprim-sulfamethoxazole (96%), and tetracycline (80%), in addition to resistance to ciprofloxacin (88%) and pipercillin-tazobactam (64%) as described previously in Europe and Asia (27,28,37). Whether such high rates of resistance are associated with the dominant spread of CTX-M-15–, rather than CTX-M-14–producing K. pneumoniae, in these patients is not known. The coexistence of CTX-M ESBL and TEM-1 and SHV-type β-lactamases in these isolates may have also contributed to the observed high rate of antimicrobial drug resistance. All except 1 of our CTX-M–positive K. pneumoniae isolates produced SHV- and CTX-M–type ESBLs. These findings have clinical implications for selecting empiric antimicrobial drug therapy when infection caused by ESBL-producing K. pneumoniae is suspected. The rapid emergence of such CTX-M–producing K. pneumoniae isolates, mainly in US hospitals, is also raising new concerns for public health and infection control practice. Third, none of the 228 KPC-producing K. pneumoniae isolates examined carried blaCTX-M. Coexistence of blaKPC and blaCTX-M has only been reported in KPC-producing K. pneumoniae in China (40). Whether certain genetic mechanisms prevent KPC-producing K. pneumoniae from acquiring blaCTX-M is unclear.
This study reveals the rapid emergence and polyclonal spread of CTX-M–producing K. pneumoniae in patients in Westchester County, New York. A limitation of our study is that the clinical isolates were collected from patients at a single tertiary-care medical center. Investigations of CTX-M–producing K. pneumoniae isolates from a variety of geographic regions should be undertaken to clarify the epidemiology and clinical and public health effects of the emergence of CTX-M–producing K. pneumoniae in the United States.
Dr. Wang is the chief of microbiology at the Westchester Medical Center and assistant professor at New York Medical College. His primary research interests include the epidemiology, mechanisms, and molecular diagnosis of emerging antimicrobial drug resistance and tick-borne diseases.
Acknowledgments
We thank Ira Schwartz and Maria Aguero-Rosenfeld for helpful advice and generous support. We are also indebted to the technologists at the Westchester Medical Center Clinical Microbiology Laboratory for saving the study isolates and for their technical contributions.
This study was supported by an intramural grant of New York Medical College (No. 46-356-1) sponsored by the Castle-Krob Foundation.
References
- Rossolini GM, D’Andrea MM, Mugnaioli C. The spread of CTX-M-type extended-spectrum beta-lactamases. Clin Microbiol Infect. 2008;14(Suppl 1):33–41. DOIPubMedGoogle Scholar
- Bonnet R. Growing group of extended-spectrum beta-lactamases: the CTX-M enzymes. Antimicrob Agents Chemother. 2004;48:1–14. DOIPubMedGoogle Scholar
- Paterson DL, Hujer KM, Hujer AM, Yeiser B, Bonomo MD, Rice LB, Extended-spectrum beta-lactamases in Klebsiella pneumoniae bloodstream isolates from seven countries: dominance and widespread prevalence of SHV- and CTX-M-type beta-lactamases. Antimicrob Agents Chemother. 2003;47:3554–60. DOIPubMedGoogle Scholar
- Moland ES, Black JA, Hossain A, Hanson ND, Thomson KS, Pottumarthy S. Discovery of CTX-M-like extended-spectrum beta-lactamases in Escherichia coli isolates from five US States. Antimicrob Agents Chemother. 2003;47:2382–3. DOIPubMedGoogle Scholar
- Lewis JS, Herrera M, Wickes B, Patterson JE, Jorgensen JH. First report of the emergence of CTX-M-type extended-spectrum beta-lactamases (ESBLs) as the predominant ESBL isolated in a U.S. health care system. Antimicrob Agents Chemother. 2007;51:4015–21. DOIPubMedGoogle Scholar
- Sidjabat HE, Paterson DL, Adams-Haduch JM, Ewan L, Pasculle AW, Muto CA, Molecular epidemiology of CTX-M-producing Escherichia coli isolates at a tertiary medical center in western Pennsylvania. Antimicrob Agents Chemother. 2009;53:4733–9. DOIPubMedGoogle Scholar
- Johnson JR, Urban C, Weissman SJ, Jorgensen JH, Lewis JS II, Hansen G, Molecular epidemiological analysis of Escherichia coli sequence type ST131 (O25:H4) and blaCTX-M-15 among extended-spectrum-beta-lactamase-producing E. coli from the United States, 2000 to 2009. Antimicrob Agents Chemother. 2012;56:2364–70. DOIPubMedGoogle Scholar
- Qi C, Pilla V, Yu JH, Reed K. Changing prevalence of Escherichia coli with CTX-M-type extended-spectrum beta-lactamases in outpatient urinary E. coli between 2003 and 2008. Diagn Microbiol Infect Dis. 2010;67:87–91. DOIPubMedGoogle Scholar
- Urban C, Mariano N, Bradford PA, Tuckman M, Segal-Maurer S, Wehbeh W, Identification of CTX-M beta-lactamases in Escherichia coli from hospitalized patients and residents of long-term care facilities. Diagn Microbiol Infect Dis. 2010;66:402–6 . DOIPubMedGoogle Scholar
- Tiruvury H, Johnson JR, Mariano N, Grenner L, Colon-Urban R, Erritouni M, Identification of CTX-M beta-lactamases among Escherichia coli from the community in New York City. Diagn Microbiol Infect Dis. 2012;72:248–52. DOIPubMedGoogle Scholar
- McGettigan SE, Hu B, Andreacchio K, Nachamkin I, Edelstein PH. Prevalence of CTX-M beta-lactamases in Philadelphia, Pennsylvania. J Clin Microbiol. 2009;47:2970–4. DOIPubMedGoogle Scholar
- Castanheira M, Sader HS, Jones RN. Antimicrobial susceptibility patterns of KPC-producing or CTX-M-producing Enterobacteriaceae. Microb Drug Resist. 2010;16:61–5. DOIPubMedGoogle Scholar
- Sjölund M, Yam J, Schwenk J, Joyce K, Medalla F, Barzilay E, Human Salmonella infection yielding CTX-M beta-lactamase, United States. Emerg Infect Dis. 2008;14:1957–9. DOIPubMedGoogle Scholar
- Folster JP, Pecic G, Krueger A, Rickert R, Burger K, Carattoli A, Identification and characterization of CTX-M-producing Shigella isolates in the United States. Antimicrob Agents Chemother. 2010;54:2269–70. DOIPubMedGoogle Scholar
- Hanson ND, Moland ES, Hong SG, Propst K, Novak DJ, Cavalieri SJ. Surveillance of community-based reservoirs reveals the presence of CTX-M, imported AmpC, and OXA-30 beta-lactamases in urine isolates of Klebsiella pneumoniae and Escherichia coli in a U.S. community. Antimicrob Agents Chemother. 2008;52:3814–6. DOIPubMedGoogle Scholar
- Lascols C, Hackel M, Hujer AM, Marshall SH, Bouchillon SK, Hoban DJ, Using nucleic acid microarrays to perform molecular epidemiology and detect novel beta-lactamases: a snapshot of extended-spectrum beta-lactamases throughout the world. J Clin Microbiol. 2012;50:1632–9 . DOIPubMedGoogle Scholar
- Castanheira M, Mendes RE, Rhomberg PR, Jones RN. Rapid emergence of blaCTX-M among Enterobacteriaceae in U.S. medical centers: molecular evaluation from the MYSTIC Program (2007). Microb Drug Resist. 2008;14:211–6. DOIPubMedGoogle Scholar
- Hoban DJ, Lascols C, Nicolle LE, Badal R, Bouchillon S, Hackel M, Antimicrobial susceptibility of Enterobacteriaceae, including molecular characterization of extended-spectrum beta-lactamase-producing species, in urinary tract isolates from hospitalized patients in North America and Europe: results from the SMART study 2009–2010. Diagn Microbiol Infect Dis. 2012;74:62–7. DOIPubMedGoogle Scholar
- Clinical Laboratory and Standards Institute. Performance standards for antimicrobial susceptibility testing: twentieth supplemental information. CLSI document M100–S20. Wayne (PA): The Institute; 2010.
- Lomaestro BM, Tobin EH, Shang W, Gootz T. The spread of Klebsiella pneumoniae carbapenemase-producing K. pneumoniae to upstate New York. Clin Infect Dis. 2006;43:e26–8. DOIPubMedGoogle Scholar
- Gröbner S, Linke D, Schutz W, Fladerer C, Madlung J, Autenrieth IB, Emergence of carbapenem-non-susceptible extended-spectrum beta-lactamase-producing Klebsiella pneumoniae isolates at the university hospital of Tubingen, Germany. J Med Microbiol. 2009;58:912–22. DOIPubMedGoogle Scholar
- Moland ES, Hanson ND, Herrera VL, Black JA, Lockhart TJ, Hossain A, Plasmid-mediated, carbapenem-hydrolysing beta-lactamase, KPC-2, in Klebsiella pneumoniae isolates. J Antimicrob Chemother. 2003;51:711–4. DOIPubMedGoogle Scholar
- Diancourt L, Passet V, Verhoef J, Grimont PA, Brisse S. Multilocus sequence typing of Klebsiella pneumoniae nosocomial isolates. J Clin Microbiol. 2005;43:4178–82. DOIPubMedGoogle Scholar
- Arlet G, Rouveau M, Casin I, Bouvet PJ, Lagrange PH, Philippon A. Molecular epidemiology of Klebsiella pneumoniae strains that produce SHV-4 beta-lactamase and which were isolated in 14 French hospitals. J Clin Microbiol. 1994;32:2553–8 .PubMedGoogle Scholar
- Johnson JR, Johnston B, Clabots C, Kuskowski MA, Castanheira M. Escherichia coli sequence type ST131 as the major cause of serious multidrug-resistant E. coli infections in the United States. Clin Infect Dis. 2010;51:286–94. DOIPubMedGoogle Scholar
- Rogers BA, Sidjabat HE, Paterson DL. Escherichia coli O25b-ST131: a pandemic, multiresistant, community-associated strain. J Antimicrob Chemother. 2011;66:1–14. DOIPubMedGoogle Scholar
- Titelman E, Iversen A, Kahlmeter G, Giske CG. Antimicrobial susceptibility to parenteral and oral agents in a largely polyclonal collection of CTX-M-14 and CTX-M-15-producing Escherichia coli and Klebsiella pneumoniae. APMIS. 2011;119:853–63. DOIPubMedGoogle Scholar
- Shin J, Kim DH, Ko KS. Comparison of CTX-M-14- and CTX-M-15-producing Escherichia coli and Klebsiella pneumoniae isolates from patients with bacteremia. J Infect. 2011;63:39–47. DOIPubMedGoogle Scholar
- Kuroda H, Yano H, Hirakata Y, Arai K, Endo S, Kanamori H, Molecular characteristics of extended-spectrum beta-lactamase-producing Escherichia coli in Japan: Emergence of CTX-M-15-producing E. coli ST131. Diagn Microbiol Infect Dis. 2012;74:201–3. DOIPubMedGoogle Scholar
- Ruiz de Alegría C, Rodriguez-Bano J, Cano ME, Hernandez-Bello JR, Calvo J, Roman E, Klebsiella pneumoniae strains producing extended-spectrum beta-lactamases in Spain: microbiological and clinical features. J Clin Microbiol. 2011;49:1134–6. DOIPubMedGoogle Scholar
- Clímaco EC, Minarini LA, da Costa Darini AL. CTX-M-producing Klebsiella spp. in a Brazilian hospital: what has changed in 6 years? Diagn Microbiol Infect Dis. 2010;68:186–9. DOIPubMedGoogle Scholar
- Pitout JD, Hossain A, Hanson ND. Phenotypic and molecular detection of CTX-M-beta-lactamases produced by Escherichia coli and Klebsiella spp. J Clin Microbiol. 2004;42:5715–21. DOIPubMedGoogle Scholar
- Wang G, Kryszuk AA, Wang K, Granada M, Schwartz I, Wormser GP. Phenotypic and molecular characterization of carbapenem-resistant Klebsiella pneumoniae clinical isolates (2005–2010). Proceedings of the 111th American Society for Microbiology General Meeting, New Orleans, Louisiana, May 21–24, 2011. Washington: American Society for Microbiology. Abstract C-604.
- Pournaras S, Protonotariou E, Voulgari E, Kristo I, Dimitroulia E, Vitti D, Clonal spread of KPC-2 carbapenemase-producing Klebsiella pneumoniae strains in Greece. J Antimicrob Chemother. 2009;64:348–52. DOIPubMedGoogle Scholar
- Sandegren L, Linkevicius M, Lytsy B, Melhus A, Andersson DI. Transfer of an Escherichia coli ST131 multiresistance cassette has created a Klebsiella pneumoniae-specific plasmid associated with a major nosocomial outbreak. J Antimicrob Chemother. 2012;67:74–83. DOIPubMedGoogle Scholar
- Karim A, Poirel L, Nagarajan S, Nordmann P. Plasmid-mediated extended-spectrum beta-lactamase (CTX-M-3 like) from India and gene association with insertion sequence ISEcp1. FEMS Microbiol Lett. 2001;201:237–41 .PubMedGoogle Scholar
- Oteo J, Cuevas O, Lopez-Rodriguez I, Banderas-Florido A, Vindel A, Perez-Vazquez M, Emergence of CTX-M-15-producing Klebsiella pneumoniae of multilocus sequence types 1, 11, 14, 17, 20, 35 and 36 as pathogens and colonizers in newborns and adults. J Antimicrob Chemother. 2009;64:524–8. DOIPubMedGoogle Scholar
- Damjanova I, Toth A, Paszti J, Hajbel-Vekony G, Jakab M, Berta J, Expansion and countrywide dissemination of ST11, ST15 and ST147 ciprofloxacin-resistant CTX-M-15-type beta-lactamase-producing Klebsiella pneumoniae epidemic clones in Hungary in 2005–the new 'MRSAs'? J Antimicrob Chemother. 2008;62:978–85. DOIPubMedGoogle Scholar
- Peirano G, Sang JH, Pitondo-Silva A, Laupland KB, Pitout JD. Molecular epidemiology of extended-spectrum-beta-lactamase-producing Klebsiella pneumoniae over a 10 year period in Calgary, Canada. [PubMed]. J Antimicrob Chemother. 2012;67:1114–20. DOIPubMedGoogle Scholar
- Chen S, Hu F, Xu X, Liu Y, Wu W, Zhu D, High prevalence of KPC-2-type carbapenemase coupled with CTX-M-type extended-spectrum beta-lactamases in carbapenem-resistant Klebsiella pneumoniae in a teaching hospital in China. Antimicrob Agents Chemother. 2011;55:2493–4. DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 19, Number 11—November 2013
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Guiqing Wang, Department of Pathology Clinical Laboratories, Westchester Medical Center, Macy Pavilion, Rm 1J-04, 100 Woods Rd, Valhalla, NY 10595, USA
Top