Volume 2, Number 3—July 1996
Synopsis
Application of Molecular Techniques to the Diagnosis of Microsporidial Infection
Cite This Article
Citation for Media
Abstract
Microsporidia are now recognized as important pathogens of AIDS patients; the ability of these parasites to cause disease in immunocompetent persons is still being elucidated. Improved diagnostic tests for microsporidial infection are continually being sought for establishing diagnosis in order to avoid laborious electron microscopy studies that require invasively acquired biopsy specimens. Modified trichrome or chemofluorescent stains are useful for detecting microsporidia in bodily fluids and stool specimens, but they do not allow for speciation of microsporidia. Polymerase chain reaction with specific primers will allow the detection and speciation of microsporidia in biopsy tissue, bodily fluids, and stool specimens.
Microsporidiosis is truly an emerging infectious disease. Although microsporidia were discovered more than 100 years ago (1), the first well-documented case of human microsporidiosis was not reported until 1959 (2). Human microsporidiosis remained an uncommon infection until the human immunodeficiency virus (HIV) pandemic; the first cases of microsporidial infection in HIV-infected patients were reported in 1985 (3,4). Since then, more than 400 HIV-associated microsporidial infections have been reported, which indicates that microsporidia are common opportunistic pathogens in patients with AIDS (5). Infections caused by three new species, Enterocytozoon bieneusi, Encephalitozoon intestinalis, and Encephalitozoon hellem, have been described in patients with acquired immunodeficiency syndrome (AIDS). Originally thought to be an opportunistic pathogen of AIDS patients exclusively, E. bieneusi was reported in 1994 to be the cause of acute, self-limited traveler's diarrhea in an immunocompetent person who was not infected with HIV (6).
As microsporidia have been increasingly recognized as pathogens of both immunosuppressed and immunocompetent persons, the need for rapid and specific diagnosis of microsporidial infection has arisen. For instance, albendazole therapy can cure E. intestinalis infections but is reported to be of little benefit to patients infected with E. bieneusi (5); therefore, identifying microsporidia to species level could have important implications in the clinical management of patients. Current diagnostic methods for microsporidia infection, though continually being improved, have shortcomings that molecular diagnostic techniques may be able to overcome.
Microsporidia are obligate intracellular parasites that infect most invertebrates and all classes of vertebrates. They belong to the phylum Microspora in the subkingdom Protozoa. They are considered ancient eukaryotes that multiply by binary fission and have a membrane bound nucleus; they lack mitochondria and Golgi membranes (5). In addition to causing disease in humans, these parasites cause disease in insects, mammals, and fish; therefore, they may create economic headaches for industries such as fisheries and silk production. They may also be beneficial to humans as biologic control agents for such pests as grasshoppers and locusts (5).
Figure 1
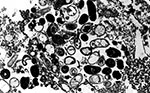
Figure 1. Transmission electron micrograph of a host cell from cell culture parasitized by Encephalitozoon intestinalis. Both vegetative forms and spores can be observed inside the parasitophorous vacuole. Original magnification, x5,200.
Figure 2

Figure 2. Transmission electron micrograph showing the coiled tubule within an Encephalitozoon intestinalis spore from cell culture. This unique structure is diagnostic of microsporidial spores. Original magnification, x28,500.
In host cells, microsporidia replicate either in a parasitophorous vacuole, like the members of the genus Encephalitozoon, or directly in the cytoplasm like E. bieneusi. Vegetative and spore stages of the organisms can be found in the host cell as the parasite undergoes merogony and sporogony, resulting in the production of the infective spore stage of the parasite (7) (Figure 1). The spores of microsporidia contain the uniquely characteristic coiled polar tubule (Figure 2). Under appropriate conditions within a suitable host, the polar tubule of the spore is extruded (Figures 3 and 4). Contact of the end of the tubule with a host cell membrane allows the spore to transfer its contents (sporoplasm) to initiate infection within the new host cell. An influx of calcium into the spore coincides with the extrusion of the polar tubule, and this mechanism may provide a target for therapy for microsporidiosis since calcium channel blockers have been shown to inhibit extrusion and the infection of host cells in vitro (8).
The human microsporidial pathogens and their clinical manifestations are shown in Table 1. The most common microsporidial disease is prolonged diarrhea with wasting caused by E. bieneusi or E. intestinalis in AIDS patients with CD4 T-cell counts below 50 cells/µl. Microsporidia have been reported in up to 39% of AIDS patients with diarrhea (9). Microsporidia may disseminate to cause systemic infection (Table 1); these organisms have been observed in urine, bile, and duodinal aspirates, as well as in ocular, sinus, bronchial, renal, hepatic, and other tissue (5). E. hellem, which primarily causes eye infections, has been exclusively found in AIDS patients. Ingestion and inhalation of spores have been suggested as likely modes of transmission for microsporidia (5,10). Biochemical, immunologic, and molecular studies performed on E. cuniculi isolates from mice, rabbits, and dogs indicate that the parasite can be classified into at least three strains (11). An evaluation of the immunologic and molecular characteristics of E. cuniculi isolated from humans and rabbits indicated that the isolates were an identical strain, thus suggesting that this microsporidion is a zoonotic parasite (12).
Transmission Electron Microscopy
Definitive diagnosis of microsporidial infection relies on observating microsporidia in biopsy tissue, bodily fluid specimens (e.g. urine, sinus aspirates, bile, cerebral spinal fluid), or stool examined by transmission electron microscopy (TEM). Microsporidia can be identified to genus or even species level on the basis of morphologic characteristics. All stages of the parasite's life cycle can be found in infected tissue, but only the spore stage can be observed in bodily fluids and stool. The size and ultrastructure of spores, particularly the configuration of the coiled tubule, distinguish E. bieneusi from the Encephalitozoon species (Figures 5 and 6). Limited success was reported when TEM was used to detect spores in bodily fluids and stool specimens to avoid obtaining specimens by invasive procedures (5). Tissue specimens are required for speciation within the Encephalitozoon genus because the meronts and sporonts, nuclear configuration, and location of replication in the host cell must be carefully studied (7,10). Problems with the use of TEM include its lack of sensitivity when performed on bodily fluids and stool, the requirement for invasive procedures to acquire biopsy specimens, and the laborious procedures for specimen preparation and examination.
Light and Fluorescence Microscopy
Histologic examination of biopsy specimens allows diagnosis of microsporidial infection but not genus or species identification of the parasites. Tissue stains used to detect microsporidia include hematoxylin and eosin, Gram, Giemsa, Warthin-Starry, and chromotrope 2R modified trichrome stains (13,14). The small size of the organisms and the lack of a noticeable tissue inflammatory response make microsporidia detection difficult (5). Once again, the invasive procedure required for obtaining specimens and the length of time needed for processing them are major drawbacks.
Figure 7

Figure 7. Spores of Encephalitozoon intestinalis in a stool specimen stained with modified trichrome stain. Note the characteristic equatorial line in one of the spores. Original magnification, x1,000.
Diagnosing microsporidial infections by light microscopy examination of noninvasively acquired specimens has been a challenge for laboratorians. Microsporidial spores are not observed in traditional ova and parasite examinations, and the parasites are generally overlooked in Gram-stained preparations of stool samples because their size, shape, and staining characteristics are similar to those of many enteric bacteria. Weber's modification of the trichrome stain has allowed more definitive identification of microsporidia by light microscopy (15). With this stain, microsporidial spores appear bright pinkish-red, and most have a distinctive diagonal or equatorial line that allows microsporidia and bacteria to be easily differentiated (Figure 7). The sensitivity and specificity of the modified trichrome stain have not been well established. When results of the modified trichrome stain performed on stool specimens and a TEM examination of duodenal biopsy specimens from HIV-infected patients were compared, E. bieneusi spores were detected in stool specimens in 9 of 9 patients with moderate or abundant parasites in tissue and in 8 (57%) of 14 stool specimens from patients whose tissue contained few parasites (16). This illustrates that microsporidial spores may not be observed in the stool of some infected patients with the modified trichrome stain.
Various chemofluorescent brighteners (such as calcofluor white and Uvitex 2B) bind to the endospore layer of microsporidia and allow spores to be detected quickly and easily in smears of specimens examined with a fluroescence microscope (5). Fluorescein-labeled polyclonal and monoclonal antibodies are being developed for the detection and speciation of microsporidia (5,17). Didier et al. (17) recently compared modified trichrome, calcofluor white, and a fouorescent polyclonal antibody stain and found that the polyclonal antibody stain was the least sensitive method for detecting microsporidia in stool, urine, and duodenal lavage specimens. They proposed screening specimens with calcofluor white and confirming positive smears by using a modified trichrome stain. However, modified trichrome and calcofluor white stains do not allow for speciation of microsporidia; therefore, speciesspecific antibodies should be used to provide definitive identifications. E.intestinalis-specific monoclonal antibodies have been used in an immunofluorescence assay for detecting spores in urine, stool, bronchial brush biopsy specimens, bronchoalvolar lavage fluid, and samples obtained from nasal swabs (18). E. bieneusi-specific antibodies may not be developed until the organism is successfully cultivated in long-term culture.
Cell Culture
Microsporidia have been isolated from a variety of specimen types and in a variety of cell lines (5). E. hellem, E. intestinalis, and Vittaforma corneae have been isolated from human specimens and maintained in continuous culture. Recently, E. bieneusi has been cultivated on a short-term (6 months) basis in vitro (19). Detecting microsporidia in infected cell cultures may take 3 to 10 weeks (19,20). Isolating microsporidia in cell culture as a means of diagnosing infection is laborious and lengthy and is prone to failure with specimens from nonsterile sites. Therefore, cell culture is not recommended as a routine laboratory technique for diagnosing microsporidiosis.
Serology
Serologic assays used to detect antibodies to microsporidia in human sera include immunofluorescence, immunoperoxidase, enzyme-linked immunosorbent assay (ELISA), and Western blot (21-23). The sensitivity and specificity of these methods for detecting antimicrosporidial antibody are not known because no comparative evaluations have been published. Studies have demonstrated increased rates of seropositivity to E. cuniculi in persons who live in tropical regions and have tropical diseases (21,24). Most notable were E. cuniculi seropositivity rates of 4.7% in patients with malaria and 9.1% in patients with schistosomiasis (24). A study of homosexual men in Sweden found that 10 (33%) of 30 were seropositive for antibodies to E. cuniculi, and all seropositive patients plus half of the seronegative patients had sometimes visited tropical countries (25). Studies to detect microsporidial antibodies in HIV-infected and non-HIV-infected patients have demonstrated that AIDS patients can mount an immune response to microsporidial infection; however, serologic methods are not useful as diagnostic tools because these studies have found that at least half of the serum specimens from persons without a history of microsporidial infection had positive titers (22,23). Some of the problems with serologic testing include the poor response to antigen challenge in immunosuppressed persons (26), the probability that different (both pathogenic and nonpathogenic) microsporidia contain common crossreactive antigens (23,26), and the lack of species-specific reagents in part because E. bieneusi cannot be grown in continuous culture.
Characterization of the microsporidial genome has focused on the small subunit ribosomal RNA (SSU-rRNA) gene. The sequence of the SSU-rRNA gene of the microsporidium Vairimorpha necatrix was published by Vossbrinck et al. in 1987 (27). The V. necatrix SSU-rRNA gene was far shorter than a typical eukaryotic SSU-rRNA gene and lacked several universal (and eukaryotic-specific) sequences. PCR amplification with primers complementary with conserved sequences of the V. necatrix SSU-rRNA gene has been used to generate sequence information from microsporidia that infect humans. SSUrRNA gene sequences have been published for E. cuniculi (28,29,30,31), E. hellem (30,31), E. intestinalis (31,32), and E. bieneusi (31,33), and an unpublished sequence for V. corneae has been deposited into GenBank National Center for Biotechnology Information, National Institutes of Health, accession 0.5 U11046.
Figure 8
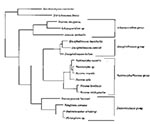
Figure 8. Cladogram representing the phylogenic relationship of several microsporidial genera as determined by small subunit ribosomal DNA sequence similarity. The human pathogens can be found in the Encephalitozoon group and the Endoreticulatus...
Taxonomic decisions concerning the microsporidia have been based historically on morphologic characteristics as established by TEM. These criteria are still valid for organisms with unique morphology and ultrastructure and were used to justify the reclassification of Nosema corneum to Vittaforma corneae (34). This reclassification was confirmed by a study of microsporidial phylogeny based on evaluation of SSU-rRNA gene sequences that indicated that N. corneum was more closely related to the insect parasite Endoreticulatus schubergi than to the other Nosema species (35) (Figure 8). Phylogenetic analysis of the sequences of the SSU-rRNA genes of microsporidia is often inconsistent with traditional classifications that are based on morphologic characteristics observed by TEM; this was demonstrated by the use of recent sequence data to determine the correct taxonomic placement of E. intestinalis (32,35). This organism was named Septata intestinalis because unique septations between spores within the parasitophorous vacuole were observable by TEM (36). Although the various stages of the microorganism throughout its life cycle are indistinguishable from those of E. cuniculi, the observation of septations allowed the organism to be identified. The E. intestinalis SSU-rRNA gene contained sufficient unique sequences to establish this organism as an independent species but shared about 90% sequence homology with the other two characterized Encephalitozoon species, E. cuniculi and E. hellem.
Molecular techniques have confirmed the existence of different strains of E. cuniculi that have been isolated from mice, rabbits, and dogs (11). Three antigenically different strains were detected by sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE) and Western blot. Double-stranded DNA heteroduplex mobility, restriction fragment length polymorphism with the restriction endonuclease Fok I, and DNA sequencing were performed on the PCR products generated by using a set of primers to amplify the entire microsporidial SSU-rRNA gene and a second set of primers specific for Encephalitozoon species. These methods allowed three strains of E. cuniculi to be clearly separated. The existence of different strains of E. bieneusi was suggested after SSU-rRNA gene sequences of organisms from maxillary sinus mucosa had been compared with those from intestinal enterocytes (37). Sequencing errors in this study were minimized by analyzing multiple recombinant DNA clones. SSU-rRNA sequences from E. bieneusi and Encephalitozoon species derived from a variety of clinical specimens must be compared to establish the existence of intraspecific genetic variation in these organisms.
Molecular Diagnosis
Primer pairs that amplify the entire microsporidial SSU-rRNA gene sequence produce amplicons of approximately 1,550 base pairs in length from Encephalitozoon species and E. bieneusi (28,33). Although these primer pairs have proven useful for sequencing and taxonomic studies, the targets are too long to be efficiently amplified from clinical specimens in a diagnostic assay. Targets for diagnostic PCR that can be amplified efficiently usually range from 100 to 400 base pairs for formalin-fixed tissue or up to 700 to 1,000 base pairs in fresh specimens (38). Several primer pairs designed to amplify short regions (250 to 607 base pairs) of the SSU-rRNA gene, and their application in the diagnosis of microsporidial infection by PCR have been published (Table 2). Primers specific for E. hellem and E. cuniculi-specific primers have been used to identify microsporidia cultured from patient specimens (39,40), but only E. bieneusi and E. intestinalis SSU-rRNA DNA have been amplified directly from patient specimens (31,33,41,42). A pair of E. bieneusi-specific primers amplified cloned E. bieneusi SSU-rRNA gene sequences but did not reliably amplify DNA from infected tissue (33). Not unexpectedly, therefore, some primer sets appear to be adequate for amplification from cultured organisms or cloned sequences but may not reliably amplify microsporidial DNA in patient specimens. The primer pair V1 and EB450 (Table 2) amplifies E. bieneusi from TEM-confirmed infected tissue and E. hellem from cell culture (33). This primer pair has not, however, been evaluated for its ability to amplify E. hellem from patient specimens. Primer pair V1 and EB450 was extensively tested by da Silva, et al. (43), who found that the primer pair did amplify E. bieneusi DNA from some patient specimens that had evidence of the parasite by electron microscopy. In addition, the primer pair did not amplify DNA of E. bieneusi derived from a short-term culture. da Silva et al. (43) described a pair of highly specific PCR primers for amplifing E. bieneusi; these are called EBIEF1 and EBIER1 (Table 2), and are based on SSU-rRNA sequences they generated. This primer pair amplified E. bieneusi DNA from cultured organisms, cloned regions of the SSU-rRNA, and patient specimens, but did not amplify SSU-rRNA-coding regions of 13 other genera and species of microsporidia. The primer pair V1 and SI500 amplifies E. intestinalis from intestinal biopsy material confirmed by TEM as infected, but does not simplify E. bieneusi-infected tissue samples or E. cuniculi from cell culture (31). This primer pair has not been evaluated for its ability to amplify other microsporidia.
Molecular techniques have confirmed the existence of different strains of E. cuniculi that have been isolated from mice, rabbits, and dogs (11). Three antigenically different strains were detected by sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE) and Western blot. Double-stranded DNA heteroduplex mobility, restriction fragment length polymorphism with the restriction endonuclease Fok I, and DNA sequencing were performed on the PCR products generated by using a set of primers to amplify the entire microsporidial SSU-rRNA gene and a second set of primers specific for Encephalitozoon species. These methods allowed three strains of E. cuniculi to be clearly separated. The existence of different strains of E. bieneusi was suggested after SSU-rRNA gene sequences of organisms from maxillary sinus mucosa had been compared with those from intestinal enterocytes (37). Sequencing errors in this study were minimized by analyzing multiple recombinant DNA clones. SSU-rRNA sequences from E. bieneusi and Encephalitozoon species derived from a variety of clinical specimens must be compared to establish the existence of intraspecific genetic variation in these organisms.
Molecular Diagnosis
Primer pairs that amplify the entire microsporidial SSU-rRNA gene sequence produce amplicons of approximately 1,550 base pairs in length from Encephalitozoon species and E. bieneusi (28,33). Although these primer pairs have proven useful for sequencing and taxonomic studies, the targets are too long to be efficiently amplified from clinical specimens in a diagnostic assay. Targets for diagnostic PCR that can be amplified efficiently usually range from 100 to 400 base pairs for formalin-fixed tissue or up to 700 to 1,000 base pairs in fresh specimens (38). Several primer pairs designed to amplify short regions (250 to 607 base pairs) of the SSU-rRNA gene, and their application in the diagnosis of microsporidial infection by PCR have been published (Table 2). Primers specific for E. hellem and E. cuniculi-specific primers have been used to identify microsporidia cultured from patient specimens (39,40), but only E. bieneusi and E. intestinalis SSU-rRNA DNA have been amplified directly from patient specimens (31,33,41,42). A pair of E. bieneusi-specific primers amplified cloned E. bieneusi SSU-rRNA gene sequences but did not reliably amplify DNA from infected tissue (33). Not unexpectedly, therefore, some primer sets appear to be adequate for amplification from cultured organisms or cloned sequences but may not reliably amplify microsporidial DNA in patient specimens. The primer pair V1 and EB450 (Table 2) amplifies E. bieneusi from TEM-confirmed infected tissue and E. hellem from cell culture (33). This primer pair has not, however, been evaluated for its ability to amplify E. hellem from patient specimens. Primer pair V1 and EB450 was extensively tested by da Silva, et al. (43), who found that the primer pair did amplify E. bieneusi DNA from some patient specimens that had evidence of the parasite by electron microscopy. In addition, the primer pair did not amplify DNA of E. bieneusi derived from a short-term culture. da Silva et al. (43) described a pair of highly specific PCR primers for amplifing E. bieneusi; these are called EBIEF1 and EBIER1 (Table 2), and are based on SSU-rRNA sequences they generated. This primer pair amplified E. bieneusi DNA from cultured organisms, cloned regions of the SSU-rRNA, and patient specimens, but did not amplify SSU-rRNA-coding regions of 13 other genera and species of microsporidia. The primer pair V1 and SI500 amplifies E. intestinalis from intestinal biopsy material confirmed by TEM as infected, but does not simplify E. bieneusi-infected tissue samples or E. cuniculi from cell culture (31). This primer pair has not been evaluated for its ability to amplify other microsporidia.
To avoid acquiring specimens by invasive procedures, PCR was used to detect microsporidia in stool specimens (42). Primer pair PMP1 and PMP2 (Table 2) allows E. bieneusi and the Encephalitozoon species (42) to be amplified and will also amplify V. corneae from culture (D.P. Fedorko, unpublished data). DNA is easily extracted from cultured organisms and biopsied tissue specimens, but extracting DNA from spores requires harsh conditions employing both mechanical and chemical disruption. A major problem with performing PCR on stool specimens is the presence of PCR inhibitors. Treating the stool specimens with sodium hypochlorite or 10% formalin before DNA extraction inactivates microorganisms in the stool and has the beneficial effect of inactivating Taq polymerase inhibitors.
Both DNA probes and restriction enzyme digestion have been used to confirm the identity of PCR amplicons (33,41,42). An internal 30 meroligonucleotide5 EB150 (5'TGTTGCGGTATTTGGTCTCTGTGTGTAAA3'), complementary to a region of the amplicon produced by primer pair V1 and EB450, has been used to detect E. bieneusi by Southern blot (31,41). Probe EB150 has been reported, however, to hybridize with E. hellem, amplified by V1 and EB450, albeit at a lower intensity than E. bieneusi (31). PCR products amplified from stool specimens by using primers PMP1 and PMP2 have been digested with restriction endonucleases HaeIII and PstI to distinguish between infection with E. bieneusi and E. intestinalis (42). These restriction enzymes do not allow E. intestinalis to be differentiated from E. cuniculi, thus limiting their use to diagnosing gastrointestinal infection.
An efficient approach for the molecular detection of microsporidia in patient specimens would involve using universal or “pan-microsporidian” primers for amplification. The species of microsporidia detected could be determined by using restriction endonuclease digestion or DNA probe assays of the amplified DNA or repeat PCR with species-specific primers. Negative specimens would require no further evaluation. Primers PMP1 and PMP2 appear to be panmicrosporidian (42), but this primer pair needs to be evaluated for its ability to amplify microsporidian DNA from a wide range of clinical specimens. Didier et al. (44) have used pan-Encephalitozoon primers that amplified a product approximately 1,000 base pairs in length, which included a large portion of the SSU-rRNA gene and a small portion of the large subunit rRNA gene. They successfully amplified E. hellem DNA from urine and conjunctival specimens from a patient with AIDS. Southern blotting and a species-specific probe were used to identify the organism to species.
The application of molecular diagnostic techniques for microsporidiosis is in its infancy. There have been no published reports of comparisons of PCR to other methods to determine sensitivity and specificity. Careful selection of primers and probes coupled with highly stringent conditions will be required to detect and speciate microsporidia in patient specimens. The potential for PCR to identify species of microsporidia from noninvasively acquired specimens makes this technique an extremely attractive diagnostic option. Although PCR can be used for detection and speciation of microsporidia in patient specimens, screening for microsporidia by using chemofluorescent stains and modified trichrome stains followed by confirmation and speciation with PCR may become the paradigm for the laboratory diagnosis of microsporidiosis.
Dr. Fedorko is a senior staff microbiologist in the Microbiology Service, Clinical Pathology Department, Warren G. Magnuson Clinical Center, National Institutes of Health, Bethesda, Maryland.
Dr. Hijazi is a senior staff pathologist in the Laboratory of Pathology, National Cancer Institute, National Institutes of Health, Bethesda, Maryland.
References
- Canning EU, Lom J, Dykova I. The microsporidia of vertebrates. New York: Academic Press, 1986.
- Matsubayashi H, Koike T, Mikata T, Hagiwara S. A case of Encephalitozoon-like body infection in man. Arch Pathol. 1959;67:181–7.
- Modigliani R, Bories C, Le Charpentier Y, Salmeron M, Messing B, Galian A, Diarrhoea and malabsorption in acquired immune deficiency syndrome: a study of four cases with special emphasis on opportunistic protozoan infections. Gut. 1985;26:179–87. DOIPubMedGoogle Scholar
- Desportes I, Le Charpentier Y, Galian A, Bernard F, CochandPriollet B, Lavergne A, . Occurrence of a new microsporidian: Enterocytozoon bieneusi n. g., n. sp., in the enterocytes of a human patient with AIDS. J Protozool. 1985;32:250–4.PubMedGoogle Scholar
- Weber R, Bryan RT, Schwartz DA, Owen RL. Human microsporidial infections. Clin Microbiol Rev. 1994;7:426–61.PubMedGoogle Scholar
- Sandfort J, Hannemann A, Gelderblom H, Stark K, Owen RL, Ruf B. Enterocytozoon bieneusi infection in an immunocompetent patient who had acute diarrhea and who was not infected with the human immuno-deficiency virus. Clin Infect Dis. 1994;19:514–6.PubMedGoogle Scholar
- Orenstein JM. Microsporidiosis in the acquired immunodeficiency syndrome. J Parasitol. 1991;77:843–64. DOIPubMedGoogle Scholar
- He Q, Leitch GJ, Visvesvara GS, Wallace S. Effects of nifedipine, metronidazole, and nitric oxide donors on spore germination and cell culture infection of the microsporidia Encephalitozoon hellem and Encephalitozoon intestinalis. Antimicrob Agents Chemother. 1996;40:179–85.PubMedGoogle Scholar
- Kotler DP, Orenstein JM. Prevalence of intestinal microsporidiosis in HIV-infected individuals referred for gastroenterological evaluation. Am J Gastroenterol. 1994;89:19982002.
- Shadduck JA, Greeley E. Microsporidia and human infections. Clin Microbiol Rev. 1989;2:158–65.PubMedGoogle Scholar
- Didier ES, Vossbrinck CR, Baker MD, Rogers LB, Bertucci DC, Shadduck JA. Identification and characterization of three Encephalitozoon cuniculi strains. Parasitology. 1995;111:411–21. DOIPubMedGoogle Scholar
- Deplazes P, Mathis A, Baumgartner R, Tanner I, Weber R. Immunologic and molecular characteristics of Encephalitozoon-like microsporidia isolated from humans and rabbits indicate that Encephalitozoon cuniculi is a zoonotic parasite. Clin Infect Dis. 1996;22:557–9.PubMedGoogle Scholar
- Kotler DP, Giang TT, Garro ML, Orenstein JM. Light microscopic diagnosis of microsporidiosis in patients with AIDS. Am J Gastroenterol. 1994;89:540–4.PubMedGoogle Scholar
- Field AS, Marriott DJ, Hing MC. The Warthin-Starry stain in the diagnosis of small intestinal microsporidiosis in HIV-infected patients. Folia Parasitol (Praha). 1993;40:261–6.PubMedGoogle Scholar
- Weber R, Bryan RT, Owen RL, Wilcox CM, Gorelkin L, Visvesvara GS. the Enteric Opportunistic Infections Working Group. Improved light-microscopic detection of microsporidia spores in stool and duodenal aspirates. N Engl J Med. 1992;326:161–6.PubMedGoogle Scholar
- Clarridge JE III, Karkhanis S, Rabeneck L, Marino B, Foote LW. Quantitative light microscopic detection of Enterocytozoon bieneusi in stool specimens: a longitudinal study of human immunodeficiency virus-infected microsporidiosis patients. J Clin Microbiol. 1996;34:520–3.PubMedGoogle Scholar
- Didier ES, Orenstein JM, Aldras A, Bertucci D, Rogers LB, Janney FA. Comparison of three staining methods for detecting microsporidia in fluids. J Clin Microbiol. 1995;33:3138–45.PubMedGoogle Scholar
- Beckers PJA, Derks GJMM, van Gool T, Rietveld FJR, Sauerwein RW. Encephalitozoon intestinalis-specific monoclonal antibodies for laboratory diagnosis of microsporidiosis. J Clin Microbiol. 1996;34:282–5.PubMedGoogle Scholar
- Visvesvara GS, Leitch GJ, Pieniazek NJ, da Silva AJ, Wallace S, Slemenda SB, Short-term in vitro culture and molecular analysis of the microsporidian, Enterocytozoon bieneusi. J Eukaryot Microbiol. 1995;42:506–10. DOIPubMedGoogle Scholar
- van Gool T, Canning EU, Gilis H, van den Bergh Weerman MA, Eeftinck Schattenkerk JKM, Dankert J. Septata intestinalis frequently isolated from stool of AIDS patients with a new cultivation method. Parasitology. 1994;109:281–9. DOIPubMedGoogle Scholar
- Hollister WS, Canning EU, Willcox A. Evidence for widespread occurence of antibodies to Encephalitozoon cuniculi (Microspora) in man provided by ELISA and other serological tests. Parasitology. 1991;102:33–43. DOIPubMedGoogle Scholar
- Didier ES, Kotler DP, Dieterich DT, Orenstein JM, Aldras AM, Davis R, Serological studies in human microsporidiosis. AIDS. 1993;7(suppl 3):S8–11.
- Didier ES, Shaddock JA, Didier PJ, Millichamp N, Vossbrinck CR. Studies on ocular microsporidia. J Protozool. 1991;38:635–8.PubMedGoogle Scholar
- Hollister WS, Canning EU. An enzyme-linked immunosorbent assay (ELISA) for detection of antibodies to Encephalitozoon cuniculi and its use in determination of infections in man. Parasitology. 1987;94:209–19. DOIPubMedGoogle Scholar
- Bergquist R, Morfeldt-Mansson L, Pehrson PO, Petrini B, Wasserman J. Antibody against Encephalitozoon cuniculi in Swedish homosexual men. Scand J Infect Dis. 1984;16:389–91. DOIPubMedGoogle Scholar
- Ombrouck C, Romestand B, Da Costa JC, Desportes-Livage I, Datry A, Coste F, Use of crossreactive antigens of the microsporidian Glugea atherinae for the possible detection of Enterocytozoon bieneusi by western blot. Am J Trop Med Hyg. 1995;52:89–93.PubMedGoogle Scholar
- Vossbrinck CR, Maddox JV, Friedman S, Debrunner-Vossbrinck BA, Woese CR. Ribosomal RNA sequence suggests microsporidia are extremely ancient eukaryotes. Nature. 1987;326:41114.
- Schuitema ARJ, Harstkeerl RA, van Gool T, Laxminarayan R, Terpstra WJ. Application of the polymerase chain reaction for the diagnosis of microsporidiosis. AIDS. 1993;7(suppl 3):S57–61.
- Hartskeerl RA, Schuitema ARJ, deWachter R. Secondary structure of the small subunit ribosomal RNA sequence of the microsporidium Encephalitozoon cuniculi. Nucleic Acids Res. 1993;21:1489. DOIPubMedGoogle Scholar
- Vossbrinck CR, Baker MD, Didier ES, Debrunner-Vossbrinck BA, Shadduck JA. Ribosomal DNA sequences of Encephalitozoon hellem and Encephalitozoon cuniculi: species identification and phylogenetic construction. J Euk Microbiol 1993;40:354-62.
- Weiss LM, Zhu X, Cali A, Tanowitz HB, Wittner M. Utility of microsporidian rRNA in diagnosis and phylogeny: a review. Folia Parasitol (Praha). 1994;41:81–90.PubMedGoogle Scholar
- Hartskeerl RA, van Gool T, Schuitema ARJ, Didier ES, Terpstra WJ. Genetic and immunological characterization of the microsporidian Septata intestinalis Cali, Kotler and Orenstein, 1993: reclassification to Encephalitozoon intestinalis. Parasitology. 1995;110:27785.
- Zhu X, Wittner M, Tanowitz HB, Kotler D, Cali A, Weiss LM. Small subunit rRNA sequence of Enterocytozoon bieneusi and its potential diagnostic role with use of the polymerase chain reaction. J Infect Dis. 1993;168:1570–5.PubMedGoogle Scholar
- Silveira H, Canning EU. Vittaforma corneae N. Comb. for the human microsporidium Nosema corneum Shadduck, Meccoli, Davis & Font, 1990 based on its ultrastructure in the liver of experimentally infected athymic mice. J Eukaryot Microbiol. 1995;42:158–65. DOIPubMedGoogle Scholar
- Baker MD, Vossbrinck CR, Didier ES, Maddox JV, Shadduck JA. Small subunit ribosomal DNA phylogeny of various microsporidia with emphasis on AIDS related forms. J Euk Microbiol 1995;42:564-70.
- Cali A, Kotler DP, Orenstein JM. Septata intestinalis, n. g., n. sp., an intestinal microsporidian associated with chronic diarrhea and dissemination in AIDS patients. J Eukaryot Microbiol. 1993;40:101–2. DOIPubMedGoogle Scholar
- Hartskeerl RA, Schuitema ARJ, van Gool T, Terpstra WJ. Genetic evidence for the occurrence of extraintestinal Enterocytozoon bieneusi infections. Nucleic Acids Res. 1993;21:4150. DOIPubMedGoogle Scholar
- Persing DH. Target selection and optimization of amplification reactions. In: Persing DH, Smith TF, Tenover FC, White TJ, editors. Diagnostic molecular microbiology: principles and applications. Washington, DC: American Society for Microbiology, 1993:88-104.
- Visvesvara GS, Leitch GJ, da Silva AJ, Croppo GP, Moura H, Wallace S, . Polyclonal and monoclonal antibody and PCR-amplified small subunit rRNA identification of a microsporidian, Encephalitozoon hellem, isolated from an AIDS patient with disseminated infection. J Clin Microbiol. 1994;32:2760–8.PubMedGoogle Scholar
- de Groote MA, Visvesvara G, Wilson ML, Pieniazek NJ, Slemenda SB, da Silva AJ, . Polymerase chain reaction and culture confirmation of disseminated Encephalitozoon cuniculi in a patient with AIDS: successful therapy with albendazole. J Infect Dis. 1995;171:1375–8.PubMedGoogle Scholar
- Franzen C, Müller A, Hegener P, Salzberger B, Hartmann P, Fätkenheuer G, . Detection of microsporidia (Enterocytozoon bieneusi) in intestinal biopsy specimens from human immunodeficiency virus-infected patients by PCR. J Clin Microbiol. 1995;33:2294–6.PubMedGoogle Scholar
- Fedorko DP, Nelson NA, Cartwright CP. Identification of microsporidia in stool specimens by using PCR and restriction endonucleases. J Clin Microbiol. 1995;33:1739–41.PubMedGoogle Scholar
- da Silva AJ, Schwartz DA, Visvesvara GS, de Moura H, Slemenda SB, Pieniazek NJ. Sensitive PCR diagnosis of infections by Enterocytozoon bieneusi (microsporidia) using primers based on the region coding for small-subunit rRNA. J Clin Microbiol. 1996;34:986–7.PubMedGoogle Scholar
- Didier ES, Rogers LB, Brush AD, Wong S, Traina-Dorge V, Bertucci D. Diagnosis of disseminated microsporidian Encephalitozoon hellem infection by PCR-Southern analysis and successful treatment with albendazole and fumagillin. J Clin Microbiol. 1996;34:947–52.PubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 2, Number 3—July 1996
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Daniel P. Fedorko, Ph.D., Clinical Pathology Department, Warren G. Magnuson Clinical Center, National Institutes of Health, Building 10, Room 2C-385, Bethesda, MD 20892-1508; fax: 301-402-1886
Top