Volume 20, Number 12—December 2014
Dispatch
Equine Influenza A(H3N8) Virus Infection in Cats
Cite This Article
Citation for Media
Abstract
Interspecies transmission of equine influenza A(H3N8) virus has resulted in establishment of a canine influenza virus. To determine if something similar could happen with cats, we experimentally infected 14 cats with the equine influenza A(H3N8) virus. All showed clinical signs, shed virus, and transmitted the virus to a contact cohort.
Equine influenza A(H3N8) virus (EIV) remains a major cause of acute respiratory infections in horses (1). Epizootics are highly explosive and spread rapidly within and among equine premises. Virus transmission is by direct contact and inhalation. First isolated in 1963, EIV has evolved and diverged into American and Eurasian lineages (2,3). The American lineage has further diverged into multiple clades: Florida-1 clade predominates in North America and Florida-2 clade in Eurasia (4).
In Florida, USA, the etiologic agent of an outbreak of acute respiratory disease among greyhounds in 2004 was identified as EIV. Virologic and serologic analyses indicated that this virus had been circulating among greyhounds for several years before. Serologic evidence of infection was also found for pet dogs (5). In Great Britain, retrospective analysis showed that an outbreak of respiratory disease among English foxhounds in 2002 was caused by an EIV (6). Likewise, this virus was found to have circulated among greyhounds in the United States before 2004 (7). The virus has now been established as canine influenza virus and has spread to other breeds and pet dogs; the virus evolved independently from EIV as a monophyletic lineage (8).
During the 2003–2004 outbreak of highly pathogenic avian influenza virus (H5N1) infection in Asia, infections in feline species, including cats, were reported. (9). Previously, given the lack of circulating feline influenza virus, feral cats had been believed to be resistant to influenza virus, although an earlier report described susceptibility to A/Hong Kong/68 (H3N2) virus infection and prolonged virus shedding (10). Recent serologic and virus isolation studies have shown that cats are susceptible to multiple influenza viruses, e.g., avian-origin canine influenza (H3N2) (11), seasonal influenza A(H1N1), and influenza A/H1N1)pdm09 (12) viruses. To investigate cats’ susceptibility to EIV infection and virus transmissibility among cats, we conducted an infection and transmission experiment.
During 2013–2014, a total of 14 specific pathogen–free domestic shorthair cats, 9–12 weeks of age, were purchased and housed in an accredited Biosafety Level 2 facility at South China Agricultural University, Guangzhou. Results of virus isolation in eggs (nasal and rectal swab samples) and serologic testing determined that these cats were influenza virus free. Experiments were approved by the Institutional Animal Care and Use Committee and monitored by veterinarians.
The virus used was A/equine/Heilongjiang/SS1/2013, which had been isolated from a mule in northern China (S. Su et al., unpub. data). For virus inoculation, 6 cats were anesthetized with xylazine hydrochloride (30 mg/kg intraperitoneally), after which they were inoculated with virus (10 TCID50 [median tissue culture infective dose]) in 1.0 mL of phosphate-buffered saline (0.5 mL in each nostril). One day after inoculation, 5 specific pathogen–free cats (contact cohort) were introduced into the same cages. Three noninfected cats (control cohort) were housed in a different room. Clinical monitoring began 1 day before virus inoculation and continued daily for the next 14 days. Nasal swab samples were collected daily for virus titration in MDCK cells. Serum was collected on postinoculation days 5, 7, 9, 12, and 14 and titrated by hemagglutination-inhibition assay with a 1% horse erythrocyte suspension. On postinoculation day 5, a total of 2 cats from the inoculated group were euthanized by intravenous pentobarbital, and on postinfection day 7, another 2 cats from the inoculated group plus 2 cats from the contact cohort were euthanized. Necropsies were performed, and trachea and lung sections were stained with hematoxylin and eosin and by an immunocytochemistry technique that involved a murine monoclonal antibody specific to EIV hemagglutinin.
Figure 1
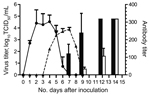
Figure 1. Results of virus titration and hemagglutination-inhibition assay for the cohort of cats inoculated with equine influenza A(H3N8) virus and the contact cohort. Virus shedding was titrated in MDCK cells. Virus titer...
The cats were susceptible to EIV infection; they showed overt clinical signs, virus shedding, and corresponding histopathologic changes in trachea and lung. Infected cats transmitted the virus to cats in the contact cohort. Overt clinical signs characteristic of acute influenza infection developed in inoculated cats during postinfection days 2–9 (peaking at day 4) and in contact cohort cats during days 4–9 (peaking at day 5); however, average clinical scores were lower for cats in the contact cohort than in the inoculated cohort (Table). Virus shedding was detected for cats in the inoculated group on days 2–5 and in the contact cohort on days 5–6 (Figure 1). This shift of virus shedding correlated with the shift in clinical signs, suggesting that the cohort group was infected by the virus shed from inoculated cats. Likewise, an antibody response was detected for cats in both groups, again 2–3 days later for the contact cohort.
Figure 2
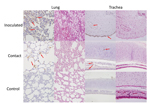
Figure 2. Detection of viral antigens in the respiratory tract of cats inoculated with equine influenza A(H3N8) virus and from a contact cohort. For each tissue type, the left column shows incubation with...
Productive viral infection was evidenced by histopathologic and immunocytochemical examinations. Characteristic lymphocytic infiltration was observed in samples from cats in the inoculated and contact cohorts; intensity was less for cats in the contact cohort (Figure 2). Likewise, EIV antigen was detected in cats in the inoculated and contact cohorts but not in the control cohort. These results indicated productive viral infection in cats in both cohorts. Because specimens were obtained from euthanized animals on the same date (postinfection day 7), the lower intensity of lymphocyte infiltration corresponded to the shift in virus titer (Figure 1) and clinical signs (Table).
That cats are susceptible to EIV by direct inoculation is not surprising because infection of cats with various influenza A viruses has been reported. Feline respiratory tract epithelial cells contain sialic acid α-2,3-galactose β-1,3-N-acetyl galactosamine (SA α2,3 gal) receptors for avian and equine influenza viruses and SA α2,6 gal receptors for mammalian influenza virus (13). However, our finding of horizontal transmission of EIV among cats is significant. If transmission occurs outside the laboratory, and if the basic reproduction rate is higher than 1.0, then EIV could potentially establish itself and circulate in this new host species. Why it has not yet happened naturally, as it did for canine influenza virus (H3N8), remains to be determined. Possibilities include lower transmission efficiency, lower probability of horse–cat contact, less virus shedding in a laboratory, or feline behavior (less social contact than dogs).
Because cats in the inoculated and contact cohorts were housed in the same cages, our study could not delineate the route of transmission (direct contact or inhalation). Experiments to elucidate the transmission mechanism are being conducted.
Because we had used a contemporary strain of EIV, to rule out the possibility that the ability to cause clinical infection is unique to this strain, we repeated the experiment with the prototype EIV, A/equine/Miami/63 (H3N8). Although 6 of 6 infected cats showed no overt clinical signs, and virus shedding was not detectable (<1:10), susceptibility was evidenced by seroconversion for 2 of the 6 inoculated cats, although at a low hemagglutination-inhibition titer (1:40 on postinoculation day 14), thereby ruling out the possibility that A/equine/Heilongjiang/SS1/2013 is an aberrant virus. None of the 3 cats in the contact cohort showed clinical signs, shed virus, or had detectable hemagglutination-inhibition titers. This strain-dependent variation in virulence is not unusual for influenza virus. Interspecies transmission of EIV to dogs and establishment of a new lineage of equine influenza virus in dogs were probably a function of that particular EIV strain, as evidenced by finding that so far only 1 EIV-originated canine influenza virus lineage is circulating. However, transmission of EIV to dogs has occasionally occurred, including during the epizootic of EIV in Australia (14). Of note, the prototype canine influenza virus is phylogenetically related to the Florida-1 clade of EIV. Whether viruses in this clade have characteristics considered to be “promiscuous” and “plastic” (15) remains to be determined. On the basis of our results, we conclude that cats are susceptible to EIV and that the infection can be transmitted by close contact.
Dr Su is a researcher at the College of Veterinary Medicine, South China Agricultural University. His research interests include the epidemiology of acute respiratory infections, influenza, and porcine viruses.
Acknowledgments
We thank Wenhua Xiang for providing EIV A/equine/Miami/63 (H3N8) virus and monoclonal antibodies for this study.
This project was supported in part by the National Natural Science Foundation of China (grant no. 31372448), the National Key Basic Research Program (project 973) of China (grant no. 2011CB504700-G), the Natural Science Foundation of Guangdong Province (no. 8251064201000008, S2012020011111), the Basic Conditions for Science and Technology Projects of Guangdong Province (no. 2011B060400015), and the Special Fund for Agro-Scientific Research in the Public Interest (grant No. 201303042).
References
- Cullinane A, Newton JR. Equine influenza—a global perspective. Vet Microbiol. 2013;167:205–14. DOIPubMedGoogle Scholar
- Daly JM, Lai AC, Binns MM, Chambers TM, Barrandeguy M, Mumford JA. Antigenic and genetic evolution of equine H3N8 influenza A viruses. J Gen Virol. 1996;77:661–71. DOIPubMedGoogle Scholar
- Lai AC, Chambers TM, Holland RE Jr, Morley PS, Haines DM, Townsend HG, Diverged evolution of recent equine-2 influenza (H3N8) viruses in the Western Hemisphere. Arch Virol. 2001;146:1063–74. DOIPubMedGoogle Scholar
- Bryant NA, Rash AS, Russell CA, Ross J, Cooke A, Bowman S, Antigenic and genetic variations in European and North American equine influenza virus strains (H3N8) isolated from 2006 to 2007. Vet Microbiol. 2009;138:41–52. DOIPubMedGoogle Scholar
- Crawford PC, Dubovi EJ, Castleman WL, Stephenson I, Gibbs EP, Chen L, Transmission of equine influenza virus to dogs. Science. 2005;310:482–5. DOIPubMedGoogle Scholar
- Daly JM, Blunden AS, Macrae S, Miller J, Bowman SJ, Kolodziejek J, Transmission of equine influenza virus to English foxhounds. Emerg Infect Dis. 2008;14:461–4. DOIPubMedGoogle Scholar
- Anderson TC, Bromfield CR, Crawford PC, Dodds WJ, Gibbs EP, Hernandez JA. Serological evidence of H3N8 canine influenza-like virus circulation in USA dogs prior to 2004. Vet J. 2012;191:312–6. DOIPubMedGoogle Scholar
- Rivailler P, Perry IA, Jang Y, Davis CT, Chen LM, Dubovi EJ, Evolution of canine and equine influenza (H3N8) viruses co-circulating between 2005 and 2008. Virology. 2010;408:71–9. DOIPubMedGoogle Scholar
- Kuiken T, Rimmelzwaan G, van Riel D, van Amerongen G, Baars M, Fouchier R, Avian H5N1 influenza in cats. Science. 2004;306:241 . DOIPubMedGoogle Scholar
- Paniker CK, Nair CM. Infection with A2 Hong Kong influenza virus in domestic cats. Bull World Health Organ. 1970;43:859–62 .PubMedGoogle Scholar
- Song DS, An DJ, Moon HJ, Yeom MJ, Jeong HY, Jeong WS, Interspecies transmission of the canine influenza H3N2 virus to domestic cats in South Korea, 2010. J Gen Virol. 2011;92:2350–5. DOIPubMedGoogle Scholar
- McCullers JA, Van De Velde LA, Schultz RD, Mitchell CG, Halford CR, Boyd KL, Seroprevalence of seasonal and pandemic influenza A viruses in domestic cats. Arch Virol. 2011;156:117–20. DOIPubMedGoogle Scholar
- Wang H, Wu X, Cheng Y, An Y, Ning Z. Tissue distribution of human and avian type sialic acid influenza virus receptors in domestic cat. Acta Vet Hung. 2013;61:537–46. DOIPubMedGoogle Scholar
- Kirkland PD, Finlaison DS, Crispe E, Hurt AC. Influenza virus transmission from horses to dogs, Australia. Emerg Infect Dis. 2010;16:699–702 .DOIPubMedGoogle Scholar
- Gibbs EP, Anderson TC. Equine and canine influenza: a review of current events. Anim Health Res Rev. 2010;11:43–51. DOIPubMedGoogle Scholar
Figures
Table
Cite This Article1These authors contributed equally to this work.
Table of Contents – Volume 20, Number 12—December 2014
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Alexander Lai, Kentucky State University, Math & Sciences, 208D Carver Hall, 400 E. Main St, Frankfort, KY 40601, USA: ; Shoujun Li, College of Veterinary Medicine, South China Agricultural University, 483 Wushan Rd, Tianhe District, Guangzhou, China 510642
Top