Volume 28, Number 12—December 2022
Research Letter
Serologic Surveillance for SARS-CoV-2 Infection among Wild Rodents, Europe
Cite This Article
Citation for Media
Abstract
We report results from serologic surveillance for exposure to SARS-CoV-2 among 1,237 wild rodents and small mammals across Europe. All samples were negative, with the possible exception of 1. Despite suspected potential for human-to-rodent spillover, no evidence of widespread SARS-CoV-2 circulation in rodent populations has been reported to date.
Esitämme tulokset serologisesta tutkimuksesta, jossa seulottiin SARS-CoV-2 tartuntojen varalta 1,237 luonnonvaraista jyrsijää ja piennisäkästä eri puolilta Eurooppaa. Kaikki näytteet olivat negatiivisia, yhtä näytettä lukuun ottamatta. SARS-CoV-2:n läikkymisen ihmisistä jyrsijöihin on arveltu olevan mahdollista, mutta todisteet viruksen laajamittaisesta leviämisestä jyrsijäpopulaatioissa puuttuvat.
Reverse transmission of diverse zoonotic pathogens (bacteria, viruses, eukaryotic parasites, fungi) from humans to animals has been recognized and documented as a global concern for years (1). On July 6, 2022, the World Organisation for Animal Health (OIE) stated, “While occasional occurrences of COVID-19 in domestic or zoo animals show little long-term consequence, infections at wildlife population levels indicate the possibility of further evolution of the virus in animals, and a future reintroduction of the virus into humans at a later date” (2). From a One Health perspective, “There is an urgent need to develop frameworks to assess the risk of SARS-CoV-2 becoming established in wild mammal populations” (3). In particular, wild rodents are suspected of being among the species more susceptible to SARS-CoV-2 infection, and susceptibility to experimental infection has been confirmed among various rodent species (4–6). Specific courses of infection may differ among rodent host species, but infection usually results in little or no detectable disease, although infectious virus may shed for 4–7 days after infection and disease may be transmitted to naive rodents (4–6). These characteristics suggest the potential for reverse transmission, broad circulation, and possible long-term establishment of SARS-CoV-2 in rodent populations. Such an event would be of concern: hamsters, for example, have transmitted SARS-CoV-2 to humans, followed by subsequent person-to-person transmission (7). Consequently, on December 6, 2021, the joint United Nations Food and Agriculture Organization and OIE (FAO-OIE) Advisory Group on SARS-CoV-2 Evolution in Animals indicated that a large surveillance study of rodent populations exposed to human contact was needed to close a major gap in SARS-CoV-2 research.
Animal experiments have shown that antibodies can be detected consistently for several weeks or longer after rodent infection with SARS-CoV-2, although detectable virus shedding lasts only a few days (4–6). When field prevalence is low or unknown among the target population, serologic testing is the preferred method to maximize chances of detecting circulation of viruses such as SARS-CoV-2 that cause brief infection but maintain longer-lasting serologic response. A recent survey in Hong Kong found a Norway rat (Rattus norvegicus) to be potentially seropositive for SARS-CoV-2 (8). Considering the high biodiversity and ubiquity of rodents, this finding called for broader surveillance studies in other continents, habitats, and noncommensal rodent species. To investigate its possible reverse zoonotic transmission and establishment in wild rodents in different settings, we conducted a large-scale serologic survey of SARS-CoV-2 in multiple rodent species across Europe.
Figure
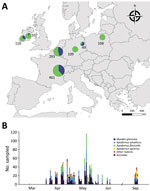
Figure. Sampling of various areas in Europe to detect SARS-CoV-2 antibody response in wild rodents. A) Location of sampling areas. Colors indicate the proportion of samples taken in the 2 habitat...
We sampled animals in urban parks and zoos, which offer ample opportunity for transmission between humans and rodents, and forests, because other wild forest mammals such as deer have become naturally infected with SARS-CoV-2 (9). During 2021, we sampled 1,202 rodents and 35 Soricidae shrews (genera Sorex and Crocidura) from 23 forests sites and 8 urban parks in 5 countries in Europe (Ireland, Belgium, France, Germany, and Poland) (Figure 1; Appendix 1 Figure 1; Appendix 2). We then assessed each rodent’s SARS-CoV-2 serologic status using an infected cell-based immunofluorescent assay (IFA; Appendix 1) (10). We chose the IFA instead of a neutralization assay as the initial screening test because it is scalable to a large number of samples and can be effective in detecting both neutralizing and nonneutralizing antibodies.
All but one of the rodents sampled were IFA negative for SARS-CoV-2. The one IFA-positive rodent (assayed twice on different days to rule out any handling error) was a wood mouse (Apodemus sylvaticus) sampled in an urban park near the city of Antwerp, Belgium, on April 6, 2021. We then tested this IFA-positive sample using a seroneutralization assay (Appendix 1), and results were negative, suggesting that the sample had no detectable neutralizing antibodies against the virus strain used in the seroneutralization assay. The sample was also negative by microsphere immunoassay (Appendix 1). The overall serologic status of this wood mouse was therefore unconfirmed. To further investigate possible virus circulation in the area, we used the Luna SARS-CoV-2 RT-qPCR Multiplex Assay Kit (New England BioLabs, https://www.neb.com) to test samples from all 59 rodents captured in the same location as the wood mouse (Appendix 1). PCRs were all negative (including for the IFA-positive wood mouse), which could be expected given the short virus-shedding period described in rodents (4–6).
Our main conclusion on the basis of this survey is that there is no evidence of a major SARS-CoV-2 spread among wild rodents in northern Europe as of April–September 2021. A similar conclusion had been reached in the study from Hong Kong (8), an area with a denser human population and large populations of pest rodents. In that study, serum from 1 urban brown rat was positive in some but not all serologic tests used, and all SARS-CoV-2 PCR tests were negative (8). Taken together, these results indicate no evidence of widespread SARS-CoV-2 circulation in rodent populations to date.
Dr. Bourret is a DVM with a PhD in virology from University of Cambridge, UK. He is a researcher at INRAE, France, and works on wildlife disease ecology and One Health topics.
Acknowledgments
We are very grateful to Jussi Hepojoki for information and advice on the IFA and Bruno Lourtet for help with Appendix Figure 1. We are also indebted to various staff at University of Helsinki, Department of Veterinary Medicine: Sofia Greilich and Akseli Valta, who helped prepare IFA slides; and Maija Suvanto and Ruut Uusitalo, who helped set up the RNA extraction protocol. We thank the animal experiment team at ANSES LRFSN for animal care and sample collection, Kalle Saksela for help with animal experiments at University of Helsinki, and Jens Jacob for supporting the project in Germany. Finally, we thank the local management teams, data management team, and land owners from Thuringia (Germany).
Our research was funded through the European H2020 (WP 2018-2020) call and the 2018–2019 BiodivERsA joint call for research proposals, under the BiodivErsA3 ERA-Net COFUND program and cofunded by Agence Nationale de la Recherche, Research Foundation–Flanders, National Science Centre, Poland, Deutsche Forschungsgemeinschaft, and the EPA Research Programme 2021–2030. The National Science Centre, Poland, supported M.G., J.N., and A.G. under the BiodivERsA3 program (2019/31/Z/NZ8/04028). Sampling from Thuringia (Germany) was funded by the DFG Priority Program 1374.
Trapping data from this study will be available in Germany in the Biodiversity Exploratories Information System (https://doi.org/10.17616/R32P9Q).
References
- Messenger AM, Barnes AN, Gray GC. Reverse zoonotic disease transmission (zooanthroponosis): a systematic review of seldom-documented human biological threats to animals. PLoS One. 2014;9:
e89055 . DOIPubMedGoogle Scholar - World Organisation for Animal Health. Crossing the species barrier: COVID-19, an example of reverse zoonosis [cited 2022 Jul 7]. https://www.woah.org/en/crossing-the-species-barriers-covid-19-an-example-of-reverse-zoonosis
- Delahay RJ, de la Fuente J, Smith GC, Sharun K, Snary EL, Flores Girón L, et al. Assessing the risks of SARS-CoV-2 in wildlife. One Health Outlook. 2021;3:7. DOIPubMedGoogle Scholar
- Bosco-Lauth AM, Root JJ, Porter SM, Walker AE, Guilbert L, Hawvermale D, et al. Peridomestic mammal susceptibility to severe acute respiratory syndrome coronavirus 2 infection. Emerg Infect Dis. 2021;27:2073–80. DOIPubMedGoogle Scholar
- Griffin BD, Chan M, Tailor N, Mendoza EJ, Leung A, Warner BM, et al. SARS-CoV-2 infection and transmission in the North American deer mouse. Nat Commun. 2021;12:3612. DOIPubMedGoogle Scholar
- Chan JFW, Zhang AJ, Yuan S, Poon VKM, Chan CCS, Lee ACY, et al. Simulation of the clinical and pathological manifestations of coronavirus disease 2019 (COVID-19) in a golden Syrian hamster model: implications for disease pathogenesis and transmissibility. Clin Infect Dis. 2020;71:2428–46. DOIPubMedGoogle Scholar
- Yen HL, Sit THC, Brackman CJ, Chuk SSY, Gu H, Tam KWS, et al.; HKU-SPH study team. Transmission of SARS-CoV-2 delta variant (AY.127) from pet hamsters to humans, leading to onward human-to-human transmission: a case study. Lancet. 2022;399:1070–8. DOIPubMedGoogle Scholar
- Miot EF, Worthington BM, Ng KH, de Lataillade LG, Pierce MP, Liao Y, et al. Surveillance of rodent pests for SARS-CoV-2 and other coronaviruses, Hong Kong. Emerg Infect Dis. 2022;28:467–70. DOIPubMedGoogle Scholar
- Kuchipudi SV, Surendran-Nair M, Ruden RM, Yon M, Nissly RH, Vandegrift KJ, et al. Multiple spillovers from humans and onward transmission of SARS-CoV-2 in white-tailed deer. Proc Natl Acad Sci U S A. 2022;119:
e2121644119 . DOIPubMedGoogle Scholar - Haveri A, Smura T, Kuivanen S, Österlund P, Hepojoki J, Ikonen N, et al. Serological and molecular findings during SARS-CoV-2 infection: the first case study in Finland, January to February 2020. Euro Surveill. 2020;25:
2000266 . DOIPubMedGoogle Scholar
Figure
Cite This ArticleOriginal Publication Date: November 02, 2022
Table of Contents – Volume 28, Number 12—December 2022
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Vincent Bourret, UR 0035 CEFS, INRAE, 24 chemin de Borde-Rouge, Auzeville CS 52627, 31326 Castanet Tolosan CEDEX, France
Top