Volume 28, Number 4—April 2022
Dispatch
Zoonotic Pathogens in Wildlife Traded in Markets for Human Consumption, Laos
Cite This Article
Citation for Media
Abstract
We tested animals from wildlife trade sites in Laos for the presence of zoonotic pathogens. Leptospira spp. were the most frequently detected infectious agents, found in 20.1% of animals. Rickettsia typhi and R. felis were also detected. These findings suggest a substantial risk for exposure through handling and consumption of wild animal meat.
Consumption of wildlife meat drives emerging infectious diseases (1), often amplified by human encroachment into natural areas and changes in land use. Wildlife trade and consumption have been responsible for outbreaks of diseases such as HIV-1 (2), Ebola (3), and monkeypox (4) and possibly for the coronavirus disease pandemic (5). Wildlife markets bring diverse species into contact, usually in dense and unsanitary conditions, enabling mixing, amplification, and transmission of pathogens among species, including humans (6). Small mammals host diverse pathogenic bacteria and viruses (7), but little investigation of endemic bacteria transmission has occurred. Determining pathogens present in traded wildlife is vital to guide appropriate measures to combat zoonotic diseases and document societal and environmental costs of wildlife trade.
Figure
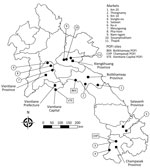
Figure. Wildlife trade sites and POFI sites (black circles) where wildlife samples were collected for study of zoonotic pathogens in wildlife traded in markets for human consumption, Laos. Provinces are labeled...
During December 2014–September 2017, we collected samples from 9 wildlife trade hotspots (8) and 2 roadside stalls (hereafter all referred to as trade sites) in Laos (Figure; Appendix Table 1). In addition, 3 Provincial Offices of Forest Inspection (POFI) collected samples from wildlife confiscated in markets by law enforcement. After identifying wildlife at trade sites (9), we asked vendors for permission to sample their animals. Depending on whether the animal was alive, dead, or butchered, we collected urogenital swabs, urine and blood samples, and kidney, liver, and spleen tissue samples (Appendix Table 2).
We extracted nucleic acid using QIAamp Viral RNA Mini Kits (QIAGEN, https://www.qiagen.com) with modifications (Appendix). We conducted PCRs targeting Leptospira spp., Rickettsia spp., Orientia tsutsugamushi, Anaplasmataceae, Ehrlichia chaffeensis, Anaplasma phagocytophilum, Coxiella burnetti, flaviviruses, hantavirus, dengue virus, Zika virus, and universal bacterial 16S rRNA (Appendix Table 3). Where necessary, PCR products were sequenced (Macrogen Inc., https://www.macrogen.com) and compared against GenBank through blastn (https://blast.ncbi.nlm.nih.gov). We performed descriptive, univariate, and multivariate analyses by using R version 3.6.2 (https://www.r-project.org). We assessed the effect of the wild meat processing status (alive, fresh, or frozen) on the risk for Leptospira detection by using a mixed effects logistic regression with species as random effect. Statistical significance was set at α = 0.05 (Appendix).
We collected 717 samples from 359 animals (trade sites: 461 samples from 324 animals; POFI: 256 samples from 35 animals); animals sampled were from >37 identifiable vertebrate species from 12 families (Appendix Table 4). Most were Sciuridae squirrels (73.0%, 262/359) and represented 16 species, most frequently Pallas’s squirrel (Callosciurus erythraeus) (20.3%, 73/359). From trade sites, 69 animals (21.3%, 95% CI 17.0%–26.2%) had >1 samples positive for >1 pathogens in 10 of 11 sites (90.9%, 95% CI 57.1%–99.5%) (Appendix Table 5). Of 324 animals tested, 65 (20.1%, 95% CI 15.9%–24.9%) were positive for Leptospira spp.; 4/41 were positive for Rickettsia spp. (9.8%, 95% CI 3.2%–24.1%), 0 for O. tsutsugamushi (0%, 95% CI 0%–10.7%), and 2 for Anaplasmataceae (4.9%, 95% CI 0.8%–17.8%) (Table 1). Positivity was higher among animals collected by POFI; 25/35 (71.4%) animals tested positive for >1 pathogens. Of those, 9 were positive for Leptospira spp. (25.7%, 95% CI 13.1%–43.6%), 20 for Rickettsia spp. (57.1%, 95% CI 39.5%–73.2%), 2 for O. tsutsugamushi (5.7%, 95% CI 1.0%–20.5%), and 6 for Anaplasmataceae (17.1%, 95% CI 7.2%–34.3%) (Table 2). Sequencing identified R. typhi, R. felis, R. conorii, an Anaplasma species (either A. centrale, A. capra, or A. marginale), A. platys, A. bovis, A. phagocytophilum, Ehrlichia chaffeensis, Lactococcus garvieae, and Kurthia populi (Tables 1, 2). No samples were positive for C. burnetii (0/76), flaviviruses (0/359), dengue virus (0/359), or Zika virus (0/358).
Among species for which >10 individual animals were sampled in trade sites, 2 had particularly high proportions of Leptospira spp.–positive specimens: the variable squirrel (Callosciurus finlaysonii) (13/28; 46.4% 95% CI 28.0%–65.8%) and the common palm civet (Paradoxurus hermaphroditus) (10/22; 45.5%, 95% CI 25.2%–67.3%). Leptospira spp.–positivity was higher in dry (50/195; 25.6%, 95% CI 19.8%–32.5%) than wet season (15/129; 11.6%, 95% CI 6.9%–18.8%) (χ2 = 8.7; p = 0.003). Data disaggregation by species and province suggested that observed seasonality was driven by results in common palm civets and variable squirrels in Champasak Province. No association was detected between the probability of an animal testing positive for Leptospira and the animal being alive (3/22; 14%, 95% CI 3.6%–36%), freshly dead (58/293; 20%, 95% CI 16%–25%; p = 0.6), or frozen (4/9; 44%, 95% CI 15%–77%; p = 0.1). In a subset of Leptospira spp.–positive animals with multiple samples, 75% (18/24; 95% CI 53%–89%) of urogenital swab samples and 50% (9/18; 95% CI 29%–71%) of blood samples were positive (p = 0.11 by Fisher exact test). Rickettsia spp. were detected exclusively in solid organs (liver, kidney, and spleen).
Zoonotic pathogens were nearly ubiquitous across sites; 10/11 sites yielded >1 pathogens. Squirrels are frequently traded in Lao markets (8) and had the greatest pathogen diversity in this study. Leptospira spp. was identified most frequently, found in 20.1% of animals (>45% in variable squirrels and common palm civets). Variable squirrels are commonly traded, often in batches of 2 to 3 squirrels (8); hence, on average, someone purchasing 3 variable squirrels would have an 83% likelihood of buying >1 infected squirrel (p = 1 – (1 – prevalence)3 = 1 – 0.553 = 0.83). The higher risk for Leptospira detection in the dry season is at odds with the typically described correlation of transmission with precipitation and flooding (10), suggesting that much remains to be understood of Leptospira ecology. Other studies have shown higher prevalence in rats (11), and although we are confident of the results from trade sites, storage of animals from POFI sites might have resulted in cross-contamination, which warrants cautious interpretation of results in this subset. Among Leptospira spp.-positive animals, detection was more likely in urogenital swab samples, highlighting the risk for transmission through infected urine (10). Although reservoir rodents are characterized by chronic renal infections, septicemia occurs during initial infection (10), and the high proportion of positive blood samples indicates a public health risk in relation to the consumption of uncooked or undercooked meat, organs, and blood. The PCR used to detect leptospires is specific for pathogenic and intermediate species (Appendix Table 3), but we could not confirm their human pathogenicity. The high volume of squirrel trade combined with high infection frequency suggests a high risk for exposure among wildlife consumers. Because leptospirosis is a key cause of fever in rural Laos (12), further work is needed to learn more about the relevance of contact with wildlife through trade and consumption.
The Rickettsiales species identified here are known to cause human infections in Laos (13). R. typhi causes murine typhus, a major underrecognized cause of fever (13). O. tsutsugamushi is responsible for up to 23% of fever (14), and although commonly associated with ground-dwelling rodents, the vectors (Leptotrombidium mites) parasitize squirrels (15), and O. tsutsugamushi has been isolated from Callosciurus notatus squirrels in Malaysia (16). Other bacteria identified are reviewed elsewhere (Appendix Table 6).
Although many of the human pathogens identified are transmitted by arthropod vectors, we found few arthropods in the wildlife sampled, probably because vectors leave animals quickly after animal death (17). Therefore, because most market vendors sell dead animals obtained from hunters or intermediaries (8), vendors are less likely to be exposed to disease vectors, and hunters are possibly at greater risk than market vendors or consumers. O. tsutsugamushi and R. typhi can cause infections through aerosol exposure, bites from infected animals, and needlestick injuries (18), but whether such routes of infection occur at trade sites is unclear. The frequent occurrence of Leptospira, which can be transmitted by direct contact with abraded skin and mucous membranes, may pose health risks to hunters, vendors, and consumers.
Dr. Nawtaisong is a postdoctoral researcher specializing in molecular microbiology of zoonotic pathogens and previously worked at the Lao-Oxford-Mahosot Hospital-Wellcome Trust Research Unit (LOMWRU).
Acknowledgments
We thank the market managers and vendors for their participation and the Provincial Offices of Forest Inspection and Provincial Livestock and Fishery Section for their assistance. We also thank the Director and staff of Mahosot Hospital and the Microbiology Laboratory, the Wildlife Conservation Society Lao PDR Program, and the Minister of Health of the Lao PDR for their support. We are very grateful to the late Rattanaphone Phetsouvanh and Manivanh Vongsouvath and Mayfong Mayxay for their support for this work.
Study protocols were reviewed and authorized by the Wildlife Conservation Society’s Institutional Animal Care and Use Committee under permit 15:04, and by the National Animal Health Laboratory, Ministry of Agriculture and Forestry, Laos.
This study was funded by the European Union under the INNOVATE program and the LACANET project (DCIASIE/2013/315-151). This work was funded in whole or in part by the Wellcome Trust (grant number: 220211). M.T.R. and P.N.N. are funded by Wellcome.
References
- Karesh WB, Cook RA, Bennett EL, Newcomb J. Wildlife trade and global disease emergence. Emerg Infect Dis. 2005;11:1000–2. DOIPubMedGoogle Scholar
- Hahn BH, Shaw GM, De Cock KM, Sharp PM. AIDS as a zoonosis: scientific and public health implications. Science. 2000;287:607–14. DOIPubMedGoogle Scholar
- Leroy EM, Rouquet P, Formenty P, Souquière S, Kilbourne A, Froment JM, et al. Multiple Ebola virus transmission events and rapid decline of central African wildlife. Science. 2004;303:387–90. DOIPubMedGoogle Scholar
- Reed KD, Melski JW, Graham MB, Regnery RL, Sotir MJ, Wegner MV, et al. The detection of monkeypox in humans in the Western Hemisphere. N Engl J Med. 2004;350:342–50. DOIPubMedGoogle Scholar
- Li X, Giorgi EE, Marichannegowda MH, Foley B, Xiao C, Kong X-P, et al. Emergence of SARS-CoV-2 through recombination and strong purifying selection. Sci Adv. 2020;6:
eabb9153 . DOIPubMedGoogle Scholar - Huong NQ, Nga NTT, Long NV, Luu BD, Latinne A, Pruvot M, et al. Coronavirus testing indicates transmission risk increases along wildlife supply chains for human consumption in Viet Nam, 2013-2014. PLoS One. 2020;15:
e0237129 . DOIPubMedGoogle Scholar - Mills JN, Childs JE. Ecologic studies of rodent reservoirs: their relevance for human health. Emerg Infect Dis. 1998;4:529–37. DOIPubMedGoogle Scholar
- Pruvot M, Khammavong K, Milavong P, Philavong C, Reinharz D, Mayxay M, et al. Toward a quantification of risks at the nexus of conservation and health: The case of bushmeat markets in Lao PDR. Sci Total Environ. 2019;676:732–45. DOIPubMedGoogle Scholar
- Francis CM. A field guide to the mammals of southeast Asia. London: New Holland; 2008.
- Blasdell KR, Morand S, Perera D, Firth C. Association of rodent-borne Leptospira spp. with urban environments in Malaysian Borneo. PLoS Negl Trop Dis. 2019;13:
e0007141 . DOIPubMedGoogle Scholar - Mayxay M, Castonguay-Vanier J, Chansamouth V, Dubot-Pérès A, Paris DH, Phetsouvanh R, et al. Causes of non-malarial fever in Laos: a prospective study. Lancet Glob Health. 2013;1:e46–54. DOIPubMedGoogle Scholar
- Phongmany S, Rolain J-M, Phetsouvanh R, Blacksell SD, Soukkhaseum V, Rasachack B, et al. Rickettsial infections and fever, Vientiane, Laos. Emerg Infect Dis. 2006;12:256–62. DOIPubMedGoogle Scholar
- Kurup A, Issac A, Loh JP, Lee TB, Chua R, Bist P, et al. Scrub typhus with sepsis and acute respiratory distress syndrome. J Clin Microbiol. 2013;51:2787–90. DOIPubMedGoogle Scholar
- Choe S, Lee D, Park H, Jeon HK, Lee Y, Na KJ, et al. Catenotaenia dendritica (Cestoda: Catenotaeniidae) and three ectoparasite species in the red squirrel, Sciurus vulgaris, from Cheongju, Korea. Korean J Parasitol. 2016;54:509–18. DOIPubMedGoogle Scholar
- Walker JS, Gan E, Muul I, Muul I; Chan Teik Chye. Involvement of small mammals in the transmission of scrub typhus in Malaysia: isolation and serological evidence. Trans R Soc Trop Med Hyg. 1973;67:838–45. DOIPubMedGoogle Scholar
- Feldhamer GA, Drickamer LC, Vessey SH, Merritt JF, Krajewski C. Mammalogy: adaptation, diversity, ecology. Baltimore (MD): Johns Hopkins University Press; 2007.
- Blacksell SD, Robinson MT, Newton PN, Day NPJ. Laboratory-acquired scrub typhus and murine typhus infections: the argument for a risk-based approach to biosafety requirements for Orientia tsutsugamushi and Rickettsia typhi laboratory activities. Clin Infect Dis. 2019;68:1413–9. DOIPubMedGoogle Scholar
Figure
Tables
Cite This ArticleOriginal Publication Date: March 09, 2022
1These first authors contributed equally to this article.
2Current affiliation: Menzies School of Health Research, Darwin, Northern Territory, Australia.
3Current affiliation: Campus Biotech, Geneva, Switzerland; University of Oxford Centre for Tropical Medicine and Global Health, Oxford, UK.
4Current affiliation: James Cook University, Townsville, Queensland, Australia.
5Current affiliation: Institut Pasteur de Madagascar, Antananarivo, Madagascar.
6These authors contributed equally to this article.
Table of Contents – Volume 28, Number 4—April 2022
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Matthew Robinson, LOMWRU, Microbiology Laboratory, Mahosot Hospital, Quai Fa Ngum, Vientiane City, Vientiane 01000, Laos
Top