Volume 28, Number 6—June 2022
Research Letter
Experimental Infection of Mink with SARS-COV-2 Omicron Variant and Subsequent Clinical Disease
Cite This Article
Citation for Media
Abstract
We report an experimental infection of American mink with SARS-CoV-2 Omicron variant and show that mink remain positive for viral RNA for days, experience clinical signs and histopathologic changes, and transmit the virus to uninfected recipients. Preparedness is crucial to avoid spread among mink and spillover to human populations.
SARS-CoV-2 has been detected in farmed and feral American mink (Neovison vison) in multiple countries, and extensive environmental contamination and human-to-mink and mink-to-human transmission has been documented (1–5). These factors have led to strict measures in mink farms and mink-farming countries to prevent the spread of the disease. In late 2021, a new SARS-CoV-2 variant (Omicron), characterized by possibly milder symptoms and more efficient human-to-human transmission, was detected, but its infectivity and spread in American mink is unknown (6,7).
We tested the response of American mink to the Omicron variant by infecting 3 male mink intranasally with 4 × 105 plaque-forming units of the virus (Appendix). We conducted follow-up on infected mink for 7 days and performed histopathologic evaluation of upper and lower respiratory tracts on the last day of follow-up. We sampled saliva daily.
All experimentally infected mink showed mild to moderate signs of illness, including lethargy, anorexia, diarrhea, nasal and lacrimal discharge, and sneezing. Consistent with earlier experiments with other variants (8; D. Adney et al., unpub. data, https://www.biorxiv.org/content/10.1101/2022.01.20.477164v1), saliva samples tested PCR-positive 1 day postinfection (dpi) and remained that way throughout follow-up (Table; Appendix). Infectious virus was cultured 1–3 dpi. Even though some of the clinical signs could be caused by other factors, such as stress from the change of environment, their consistency with signs seen in studies of other variants, combined with PCR results, demonstrate that the Omicron variant also causes clinical disease in mink.
To study whether mink can transmit the virus, we placed 2 uninfected indirect contact mink in separate cages 10–20 cm from the cages of the infected mink and followed their progress for 10 days. Similar signs to the experimentally infected mink developed in both initially uninfected mink, and they were consistently PCR-positive from day 3 onward (Table), indicating mink-to-mink transmission. Infectious virus was detected in cell culture even before it was detected by PCR. Even though no evidence of mink-to-human transmission of the Omicron variant exists, it seems possible on the basis of our results and the information from studies of other variants.
Figure
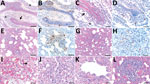
Figure. Histopathologic changes and SARS-CoV-2 expression in the upper and lower respiratory tracts of mink experimentally infected with Omicron variant at 7 days postinfection and recipient mink after 10 days of...
Gross findings in the nasal cavity and lungs were subtle in both experimentally infected and recipient mink and consisted of hyperemia of respiratory mucosa with small amounts of viscous exudate and noncollapsed, dark-red, and wet pulmonary lobes. All mink showed histopathologic changes in the upper and lower respiratory tracts (Figure). We observed multifocal degeneration and loss of respiratory epithelium with variable mucosal and submucosal neutrophilic infiltration in the nose. The lumen contained sloughed epithelial cells, mucinous material, and degenerated neutrophils (Figure, panels A, C). Viral nucleoprotein was widely distributed beyond intact cells, within sloughed cells, and in mucosal respiratory epithelium (Figure, panels B, D). The olfactory epithelium was inconsistent and mildly affected with only focal viral antigen detection. Unlike in some experimental infections reported in rodents, clear pathology was observed in the lungs (9). In 2 inoculated and both recipient mink, pulmonary lesions (Figure, panels E, G) were associated with viral antigen expression (Figure, panels F, H) and characterized by multifocal to coalescing alveolar damage with degeneration or necrosis of alveolar septa, infrequent hyalin membrane formation, and variable proliferation of type II pneumocytes (Figure, panels I, J). Alveolar spaces contained macrophages, sloughed cells, edema, and hemorrhage. Bronchiolar epithelial degeneration and hyperplasia were variably present (Figure, panel K), and the lumen filled with few sloughed cells and neutrophils. Bronchi were lined by hyperplastic epithelium with increased numbers of goblet cells. Other consistent findings were vasculitis (Figure, panel L), perivasculitis, and perivascular and peribronchial edema. In 1 inoculated mink, we observed markedly thickened alveolar septa by mononuclear cells, marked proliferation of type II pneumocytes, intra-alveolar macrophages, few syncytial cells, bronchi and bronchiolar epithelial cell hyperplasia, vasculitis, and perivasculitis. We could not detect viral antigen in this mink. Strikingly, all evaluated mink lacked viral antigen in the epithelium of bronchi and bronchioles.
The Omicron variant is different from other variants in its more efficient spread, primarily attributable to immune escape and likely milder symptoms in humans (6,7). These factors make preventing virus introduction into mink farms through asymptomatic humans more difficult, creating a more substantial risk for the formation of virus reservoirs among farmed or feral mink. This study shows that mink can be infected by Omicron and, crucially, efficiently transmit the virus to other mink. Despite the reports of lower virulence of Omicron, mink experience clinical disease and nasal and pulmonary microscopic lesions that closely resemble infection with previously reported variants in mink and humans (8). Clarifying the clinical signs will help detect the virus among mink earlier. Questions remain about the risks that the spread of this easily transmitted variant among mink would create for public health, including transmission to humans and emergence of mink-specific mutations, followed by their spillover to human population.
This article was preprinted at https://www.biorxiv.org/content/10.1101/2022.02.16.480524v2.
Ms. Virtanen is a PhD student at the University of Helsinki in the field of clinical microbiology. Her research interests include zoonotic viruses and pathogens in human-animal interface.
Acknowledgments
We thank Jari Elemo, Mari Elemo, and other animal caretakers for handling the animals and assessing their health and Esa Pohjalainen, Sanna Mäki, Tiina Sihvonen, Johanna Rintamäki, Hanna Valtonen, Marika Skön, Larissa Laine, and Elina Väisänen for technical assistance. We thank Kati Kuipers, Anne Kujanpää, Laura Vähälä, and the Finnish Centre for Laboratory Animal Pathology (FCLAP) for expert technical help, as well as Johanna Korpela and Jussi Peura from Finnish Fur Breeders Association and Jan Segervall and Maarit Mohaibes from the Kannus Research Farm Luova Ltd. for providing the animals. We also thank E3 Excellence in Pandemic Response and Enterprise Solutions coinnovation project and all its parties.
This study was supported by the Academy of Finland (grant no. 336490, 339510), VEO–European Union’s Horizon 2020 (grant no. 874735), Business Finland E3 (4917/31/2021), Finnish Institute for Health and Welfare, and the Jane and Aatos Erkko Foundation.
References
- Aguiló-Gisbert J, Padilla-Blanco M, Lizana V, Maiques E, Muñoz-Baquero M, Chillida-Martínez E, et al. First description of SARS-CoV-2 infection in two feral American mink (Neovison vison) caught in the wild. Animals (Basel). 2021;11:1422. DOIPubMedGoogle Scholar
- Oreshkova N, Molenaar RJ, Vreman S, Harders F, Oude Munnink BB, Hakze-van der Honing RW, et al. SARS-CoV-2 infection in farmed minks, the Netherlands, April and May 2020. [Erratum in: Euro Surveill. 2021;26:210325c]. Euro Surveill. 2020;25:25. DOIPubMedGoogle Scholar
- Rabalski L, Kosinski M, Mazur-Panasiuk N, Szewczyk B, Bienkowska-Szewczyk K, Kant R, et al. Zoonotic spill-over of SARS-CoV-2: mink-adapted virus in humans. Clin Microbiol Infect. 2021.PubMedGoogle Scholar
- Rabalski L, Kosinski M, Smura T, Aaltonen K, Kant R, Sironen T, et al. Severe acute respiratory syndrome coronavirus 2 in farmed mink (Neovison vison), Poland. Emerg Infect Dis. 2021;27:2333–9. DOIPubMedGoogle Scholar
- Hammer AS, Quaade ML, Rasmussen TB, Fonager J, Rasmussen M, Mundbjerg K, et al. SARS-CoV-2 transmission between mink (Neovison vison) and humans, Denmark. Emerg Infect Dis. 2021;27:547–51. DOIPubMedGoogle Scholar
- Viana R, Moyo S, Amoako DG, Tegally H, Scheepers C, Althaus CL, et al. Rapid epidemic expansion of the SARS-CoV-2 Omicron variant in southern Africa. Nature. 2022;603:679–86. DOIPubMedGoogle Scholar
- Wolter N, Jassat W, Walaza S, Welch R, Moultrie H, Groome M, et al. Early assessment of the clinical severity of the SARS-CoV-2 omicron variant in South Africa: a data linkage study. Lancet. 2022;399:437–46. DOIPubMedGoogle Scholar
- Song Z, Bao L, Deng W, Liu J, Ren E, Lv Q, et al. Integrated histopathological, lipidomic, and metabolomic profiles reveal mink is a useful animal model to mimic the pathogenicity of severe COVID-19 patients. Signal Transduct Target Ther. 2022;7:29. DOIPubMedGoogle Scholar
- Halfmann PJ, Iida S, Iwatsuki-Horimoto K, Maemura T, Kiso M, Scheaffer SM, et al.; Consortium Mount Sinai Pathogen Surveillance (PSP) study group. SARS-CoV-2 Omicron virus causes attenuated disease in mice and hamsters. Nature. 2022;603:687–92. DOIPubMedGoogle Scholar
Figure
Table
Cite This ArticleOriginal Publication Date: May 13, 2022
1These authors contributed equally to this article.
Table of Contents – Volume 28, Number 6—June 2022
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Jenni Virtanen, Department of Veterinary Biosciences, Faculty of Veterinary Medicine, University of Helsinki, Agnes Sjöbergin katu 2, 00790, Helsinki, Finland
Top