Volume 30, Number 6—June 2024
Dispatch
Concurrent Infection with Clade 2.3.4.4b Highly Pathogenic Avian Influenza H5N6 and H5N1 Viruses, South Korea, 2023
Cite This Article
Citation for Media
Abstract
Highly pathogenic avian influenza H5N6 and H5N1 viruses of clade 2.3.4.4b were simultaneously introduced into South Korea at the end of 2023. An outbreak at a broiler duck farm consisted of concurrent infection by both viruses. Sharing genetic information and international surveillance of such viruses in wild birds and poultry is critical.
Since clade 2.3.4.4 H5Nx highly pathogenic avian influenza (HPAI) viruses first emerged in East Asia in 2013‒14, clade 2.3.4.4b has spread throughout Europe, Africa, and Middle East in 2016‒17, causing >2,000 outbreaks in poultry and wild birds in >30 countries (1,2). Clade 2.3.4.4b H5N1 viruses were detected in Europe in late 2020; that clade became the predominant subtype in Europe in 2021 and spread throughout Asia and North America. By the end of 2023, H5N1 viruses of that clade had affected bird populations in most of the United States and spread to South America and Antarctica (3).
Since early 2014, South Korea has experienced epidemic outbreaks of different subtypes of this clade, including H5N8 in early 2014, 2016‒17, and 2020‒21; H5N6 in 2017‒18; and H5N1 in 2021‒22 and 2022‒23 (4–7). All those outbreaks in poultry have occurred during the winter season, when migratory birds enter and stay on the Korean peninsula (8–10). No HPAI virus was detected during regular active surveillance of both wild birds and poultry during May–October 2023.
The first suspected case of HPAI in poultry in the 2023‒24 winter season was reported in South Korea. Surprisingly, birds at that farm were found to be concurrently infected with H5N6 and H5N1 viruses of clade 2.3.4.4b. Subsequently, birds at poultry farms as well as wild birds were found to be infected with H5N6 or H5N1 viruses. Our study analyzed whole-genome sequences of the virus populations of pooled swab samples from the flocks at the farm that were infected with both H5N6 and H5N1 influenza viruses; we defined the farm as the index case. We compared those sequences with the sequences of viruses isolated from other affected farms and wild birds to determine the origins of the viruses and their relationships.
Figure 1
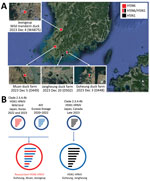
Figure 1. Locations of duck farms and of wild ducks infected with clade 2.3.4.4b HPAIV H5, South Korea, 2023, and their viral genotypes. A) Each square indicates the location of the infected...
On December 3, 2023, a suspected HPAI infection that caused white diarrhea, reduced feed intake, and increased deaths was reported in 39-day-old broiler ducks at a broiler duck farm (D448) in Goheung, South Korea (Figure 1). We detected matrix and H5 genes in the clinical samples from this index farm by real-time reverse transcription PCR. We determined the deduced amino acid sequence of the HA cleavage site of the H5 genes to be PLREKRRKR/GLF, which indicated high pathogenicity. For the NA gene, we detected both N1 and N6 genes in some flocks at the farm, at which the flocks were separated in different houses. We analyzed co-infection status at that farm by using whole-genome sequences of avian influenza viruses obtained from pooled oropharyngeal swab samples of 20 live ducks from each of 11 flocks using the Nanopore (Oxford Nanopore, https://nanoporetech.com) amplicon sequencing method (Appendix).
Figure 2

Figure 2. Co-infection status of birds at a broiler duck farm (D448) infected with clade 2.3.4.4b highly pathogenic avian influenza H5N6 and H5N1 viruses in South Korea, December 2023. Reference gene segments...
We found that 3 flocks, numbers 1, 4, and 5, were co-infected with H5N6 and H5N1 viruses, whereas the other 8 flocks were infected with H5N6 virus only (Figure 2). Analysis of the average coverage at each gene segment as percentage composition showed that birds in flock 4 had more viral reads of H5N1, whereas flocks 1 and 5 had more reads of H5N6 (Figure 2, panel B). We observed the same co-infection pattern in pooled cloacal swabs of flock 4 (data not shown). Because all the swabs from flocks were pooled at sample collection, no clear evidence was found supporting infection with the 2 viruses in a single bird. Because this farm was located very close to the south sea and seawall lake and had a relatively low level of biosecurity, we considered this farm susceptible to virus introduction by migratory birds (Figure 1, panel A).
We detected HPAI H5N6 virus (WA875) in Jeolla-do province in an apparently healthy wild mandarin duck, which we captured and sampled on December 4, 2023, for the active wild bird surveillance program. Two additional broiler-duck farms in the same province were found to be infected with H5N6 (D449) and H5N1 (D502) virus on December 5 and December 20, 2023 (Figure 1, panel A). We assessed the genetic relationships among the HPAI viruses by determining and comparing the complete genome sequences of A/duck/Korea/D448-N6/2023(H5N6), A/duck/Korea/D448-N1/2023(H5N1), A/duck/Korea/D449/2023(H5N6), A/mandarin duck/Korea/WA875/2023(H5N6), and A/duck/Korea/D502/2023(H5N1). Their sequences have been deposited in GISAID (https://www.gisaid.org; accession nos. EPI_ISL_18819959–61, EPI_ISL_18819826, and EPI_ISL_18819797).
The H5N6 viruses, D448-N6, D449, and WA875, showed high nucleotide sequence identities in all 8 genes among them (>99.8%). The sequences of their polymerase basic (PB) 1, hemagglutinin (HA), and matrix (M) genes were very close (99.53%–99.83%) to the respective genes of clade 2.3.4.4b HPAI H5N1 viruses isolated from wild birds in Japan and South Korea in 2022‒23. The 4 internal genes of the H5N6 viruses, PB2, polymerase acidic protein (PA), nucleoprotein (NP), and nonstructural protein (NS), were closely related to the respective genes found in the Eurasian low pathogenicity avian influenza (LPAI) viruses of diverse subtypes isolated from wild birds in 2020 and 2022. Their N6 genes appeared to be close to the poultry viruses isolated in East Asia in 2021 and 2022, although the nucleotide identities were relatively low (98.1%–98.62%) (Table 1). Of interest, the protein encoded by the N6 gene in the isolates from this study had a deletion of 12 aa residues at positions 58–69; this neuraminidase (NA) stalk deletion has been often observed in poultry-adapted viruses (11,12). From the avian influenza active surveillance program in South Korea during 2019–2023, N6 genes were detected only in LPAI viruses isolated from wild birds; we did not observe this NA stalk deletion among the analyzed viruses (data not shown).
We found no HPAI H5N6 viruses showing nucleotide similarities >98.5% in all of the 8 genes at once to the new H5N6 isolates in the public databases. However, a wild bird isolate from Japan (A/peregrine falcon/Saga/4112A002/2023, EPI_ISL_18740267) that was collected on December 6, 2023, was almost identical to the H5N6 Korean viruses (T. Hiono, pers. comm., email, 2024 Jan 11), suggesting that these emerged viruses spread coincidently throughout this winter in East Asia.
The nucleotide sequences of the coding regions of 2 poultry H5N1 viruses, D448-N1 and D502, were very similar (>99.0%) and were very closely related to the sequences of clade 2.3.4.4b H5N1 viruses circulating in Japan and Canada in 2023 (Table 2; Appendix Figures 1–5,7–9). Those clade 2.3.4.4b HPAI H5N1 viruses of diverse genotypes have been prevalent in Europe and North America (3) and had been introduced into South Korea during the epidemics of 2021–22 and 2022–23 (5,8). We did not detect significant mutations related to mammal adaptation or antiviral resistance in the newly isolated H5N6 and H5N1 HPAI viruses.
This study describes the simultaneous introduction of H5N1 virus and a new reassortant H5N6 HPAI virus of clade 2.3.4.4b into South Korea in 2023. Better understanding of this spatial and genomic dynamic requires enhanced and timely sharing of genetic information and international surveillance of HPAI and LPAI viruses in wild birds and poultry.
Dr. Heo is a researcher at the Animal and Plant Quarantine Agency, South Korea. His research interests include surveillance of zoonotic viruses.
Acknowledgments
We thank Jeong-Eui Lee, Byeong-Suk Jeon, and Chae-Rin Lee for excellent technical assistance. We also thank the Animal and Plant Quarantine Agency, Ministry of Agriculture, Food and Rural Affairs, and the regional office for Animal Disease Control, Livestock Health Control Association, for their efforts to control AIV. We thank Professor Takahiro Hiono of Hokkaido University of Japan for his prompt communications and colleagues worldwide for their laboratory contributions, made available through GISAID. We also thank the SDSC’s CIPRES Science Gateway at the University of California-San Diego for computing resources.
This research was supported by a grant from the Animal and Plant Quarantine Agency(B-1543418-2023-23-01) of South Korea.
Author contributions: All authors have approved the final article.
References
- Napp S, Majó N, Sánchez-Gónzalez R, Vergara-Alert J. Emergence and spread of highly pathogenic avian influenza A(H5N8) in Europe in 2016-2017. Transbound Emerg Dis. 2018;65:1217–26. DOIPubMedGoogle Scholar
- Lee DH, Bertran K, Kwon JH, Swayne DE. Evolution, global spread, and pathogenicity of highly pathogenic avian influenza H5Nx clade 2.3.4.4. J Vet Sci. 2017;18(S1):269–80. DOIPubMedGoogle Scholar
- Adlhoch C, Fusaro A, Gonzales JL, Kuiken T, Mirinavičiūtė G, Niqueux É, et al.; European Food Safety Authority; European Centre for Disease Prevention and Control; European Union Reference Laboratory for Avian Influenza. Avian influenza overview June-September 2023. EFSA J. 2023;21:
e08328 .PubMedGoogle Scholar - Seo YR, Cho AY, Si YJ, Lee SI, Kim DJ, Jeong H, et al. Evolution and spread of highly pathogenic avian influenza A(H5N1) clade 2.3.4.4b virus in wild birds, South Korea, 2022–2023. Emerg Infect Dis. 2024;30:299–309. DOIPubMedGoogle Scholar
- Kang YM, Heo GB, An SH, Lee YN, Cha RM, Cho HK, et al. Introduction of multiple novel high pathogenicity avian influenza (H5N1) virus of clade 2.3.4.4b into South Korea in 2022. Transbound Emerg Dis. 2023;2023:
8339427 . DOIGoogle Scholar - Baek YG, Lee YN, Lee DH, Cheon SH, Kye SJ, Park YR, et al. A novel reassortant clade 2.3.4.4 highly pathogenic avian influenza H5N6 virus identified in South Korea in 2018. Infect Genet Evol. 2020;78:
104056 . DOIPubMedGoogle Scholar - Lee EK, Kang HM, Song BM, Lee YN, Heo GB, Lee HS, et al. Surveillance of avian influenza viruses in South Korea between 2012 and 2014. Virol J. 2017;14:54. DOIPubMedGoogle Scholar
- Sagong M, Lee YN, Song S, Cha RM, Lee EK, Kang YM, et al. Emergence of clade 2.3.4.4b novel reassortant H5N1 high pathogenicity avian influenza virus in South Korea during late 2021. Transbound Emerg Dis. 2022;69:e3255–60. DOIPubMedGoogle Scholar
- Lee YJ, Kang HM, Lee EK, Song BM, Jeong J, Kwon YK, et al. Novel reassortant influenza A(H5N8) viruses, South Korea, 2014. Emerg Infect Dis. 2014;20:1087–9. DOIPubMedGoogle Scholar
- Nabeshima K, Takadate Y, Soda K, Hiono T, Isoda N, Sakoda Y, et al. Detection of H5N1 high pathogenicity avian influenza viruses in four raptors and two geese in Japan in the fall of 2022. Viruses. 2023;15:1865. DOIPubMedGoogle Scholar
- Li J, Zu Dohna H, Cardona CJ, Miller J, Carpenter TE. Emergence and genetic variation of neuraminidase stalk deletions in avian influenza viruses. PLoS One. 2011;6:
e14722 . DOIPubMedGoogle Scholar - Yang L, Zhu W, Li X, Bo H, Zhang Y, Zou S, et al. Genesis and dissemination of highly pathogenic H5N6 avian influenza viruses. J Virol. 2017;91:e02199–16. DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleOriginal Publication Date: May 03, 2024
Table of Contents – Volume 30, Number 6—June 2024
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Kwang-Nyeong Lee, Avian Influenza Research and Diagnostic Division, Animal and Plant Quarantine Agency, 177 Hyeoksin 8-ro, Gimcheon-si, Gyeongsangbuk-do 39660, South Korea
Top