Volume 10, Number 9—September 2004
Research
Experimental Infection of Ground Squirrels (Spermophilus tridecemlineatus) with Monkeypox Virus
Cite This Article
Citation for Media
Abstract
A proposed new small-animal (rodent) model for studying the pathogenesis and treatment of severe orthopoxvirus infections is described. Thirteen-lined ground squirrels (Spermophilus tridecemlineatus) were infected intraperitoneally and intranasally with monkeypox virus (MPXV). A fulminant illness developed in all animals, and they died 6–9 days after infection. Virus was cultured from the blood and oropharynx several days before death; at necropsy, all of the organs tested contained relatively high titers of MPXV. The major pathologic findings were in the liver, which showed centrilobular necrosis, steatosis, and basophilic inclusion bodies in hepatocytes. Splenic necrosis was also observed, as well as interstitial inflammation in the lungs. The pathologic features of MPXV in ground squirrels is similar to that described with MPXV in macaques and severe variola (smallpox) virus infection in humans.
Until last year, human monkeypox was confined to forested areas of central and West Africa, where sporadic epizootics have occurred (1). However, in 2003 monkeypox appeared in the United States, and 32 human cases were confirmed during an outbreak that occurred in pet owners in the Midwest (2,3). Imported African rodents were implicated as the probable source of the outbreak, although the virus also infected other wild animal pets (i.e., prairie dogs) that had contact with them (4). On the basis of these reports and earlier studies in Africa (5–7) that suggest that squirrels and certain other wild rodents might be reservoirs of monkeypox virus (MPXV), we tested the susceptibility of several North American wild rodent species to MPXV infection. We report the results of our studies with the common thirteen-lined ground squirrel, Spermophilus tridecemlineatus.
Animals
Ten adult thirteen-lined ground squirrels (S. tridecemlineatus) were used in the experiment. The animals were wild-caught and purchased from a commercial supplier (TLS Research, Bloomington, IL). Ground squirrels were housed individually in filter-bonneted, solid bottom (123-cm2 floor area) plastic cages in an isolation room within an animal biosafety level 3 facility. All persons handling the animals had recently received smallpox (vaccinia) vaccination and used appropriate personal protection. Animals were cared for in accordance with the guidelines of the Committee on Care and Use of Laboratory Animals (Institute of Laboratory Animal Resources, National Research Council) under an animal use protocol approved by the Institutional Animal Care and Use Committee at the University of Texas Medical Branch.
Virus
The strain of MPXV used was designated MPX 2003 and was provided by the Centers for Disease Control and Prevention, Atlanta, Georgia. This virus was originally isolated from a skin lesion from a human monkeypox patient during the 2003 U.S. outbreak (3). A stock of the virus was prepared from infected Vero cells; the unsonicated frozen cell lysate was used to infect the rodents and had a titer of 106.l PFU/mL.
Virus Assay
Samples for virus assay were stored at –80°C. Before testing, tissue samples were thawed and triturated in sterile Ten Broeck glass tissue grinders in phosphate-buffered saline (PBS), pH 7.4, containing 30% heat-inactivated (56°C for 30 min) fetal bovine serum (FBS) to prepare an approximate 10% (wt/vol) tissue homogenate. After centrifugation at 6,000 rpm for 5 min to clarify the suspension, serial 10-fold dilutions from 10–1 to 10–8 were prepared in PBS containing 10% FBS. Similar dilutions were made with the blood and throat swab suspensions for virus assay.
Dilutions of the tissue homogenates, blood, and throat swab suspensions were titrated in 24-well cultures of Vero cells; four wells were used for each dilution, as described (8). Cultures were incubated at 37°C, and plaques were counted 4–6 days later. Virus titers were defined as the number of PFU per milliliter of sample.
Experimental Infection of Animals
Ground squirrels were infected by the intraperitoneal (IP) or intranasal (IN) routes. Five squirrels were injected IP with 105.1 PFU of MPX 2003 virus. Five other animals were infected by the IN route; under Halothane (Halocarbon Laboratories, River Edge, NJ) anesthesia, two drops of the stock virus solution containing 106.1 PFU/mL were instilled into each nostril. After infection, all rodents were observed daily for signs of illness; if an animal died, a necropsy was performed, and tissues (liver, spleen, kidney, adrenal, lung, heart, and brain) were taken for histopathology and virus titration. In some animals, enlarged mesenteric lymph nodes and thymus were also taken. Blood (100 μL from the retroorbital sinus) and an oropharyngeal swab were also taken daily from each animal for virus assay. The whole blood and the swab were expressed in 900 μL of PBS with 10% FBS.
Histopathologic and Immunohistochemical Methods
At necropsy, tissue samples were taken from the animals and preserved in 10% buffered formalin for 24 to 48 h, followed by storage in 70% ethanol. After fixation, the samples were processed for routine embedding in paraffin. Four- to 5-μm–thick tissue sections were made and stained by the hematoxylin and eosin method (9).
Selected tissue sections were also studied immunohistochemically, by using a vaccinia hyperimmune mouse ascetic fluid, at a dilution of 1:100. A mouse-on-mouse IHC-ISO labeling kit (InnoGenex, San Ramon, CA) was used, according to the manufacture’s instructions and a protocol similar to one described (9). The primary antibody (a mouse antivaccinia ascitic fluid) was incubated with the section at 4°C for overnight. Tissue sections from two uninfected animals were used as negative controls.
Transmission Electron Microscopy (EM)
For EM, formalin-fixed tissues were additionally fixed in a mixture of 2.5% formaldehyde, 0.1% glutaraldehyde, 0.03% trinitrophenol, and 0.03% CaCl2 in 0.05 mol/L cacodylate buffer, postfixed in 1% OsO4 in 0.1 mol/L cacodylate buffer, stained en bloc in 1% uranyl acetate in 0.1 mol/L maleate buffer, dehydrated in ethanol, and embedded in Poly/Bed 812 (Polysciences, Warrington, PA). Ultrathin sections were cut on a Reichert-Leica Ultracut S ultramicrotome and examined in a Philips 201 electron microscope at 60 kV.
Clinical Manifestations
Most of the animals became lethargic and anorexic within 4 or 5 days after infection; however, detectable skin lesions, respiratory distress, or other obvious symptoms of disease did not develop in any of the ground squirrels. The five animals (numbers 11–15) infected IP were moribund or dead within 6 or 7 days after infection; death occurred in the IN-infected group (animals 16–20) approximately 2 days later (Table 1). All of the animals were dead within 9 days after infection.
Virus Titrations
Table 1 shows the amount of virus detected in daily blood and throat swab samples taken from the infected rodents. Among the IP-infected squirrels, virus was first detected in the blood on day 3; in contrast, in the IN-infected animals, virus was first detected in the oropharynx (throat swab) on days 2 to 4. Virus titers in the two groups of animals were similar and tended to increase with time.
Table 2 gives the results of virus titrations performed on 10% suspensions of liver, spleen, kidney, lung, heart, and brain taken at necropsy from infected animals. The highest MPXV titers were found in the liver and spleen, but relatively large amounts of virus were detected in the other organs as well. The amount of virus present in the various organ groups (i.e., liver or lung) did not appear to be related to the route of infection, a finding that suggests that the MPXV infection in the rodents was disseminated. The lower virus titers in brain and heart, compared to blood, suggest that MPXV did not replicate appreciably in those organs.
Pathologic Changes and Immunohistochemical Analysis
Figure 1
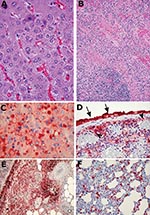
Figure 1. Representative photomicrographs of histologic changes and immunohistochemical staining of tissues from ground squirrels infected with monkeypox virus. A) Liver from a ground squirrel (intranasal infection) showing mild degenerative changes, including early...
In the IP-infected animals (numbers 11–15), considerable centrilobular hepatocytic degeneration or necrosis occurred in the liver. Many hepatocytes, particularly in the areas of degeneration, contained round to oval-shaped basophilic inclusion bodies of various sizes in the cytoplasm. Inflammatory cell infiltration in the lobules was minimal. The portal tracts were normal. Moderate-to-marked necrosis of the spleen was also present in all the animals (Figure 1B). This necrosis was characterized by lymphocytic karyorrhexis in the white pulp, and fibrinoid necrosis, congestion, and endothelial cell swelling in the red pulp, accompanied by cell debris. The lungs showed mild-to-multifocal thickening of the alveolar septa and focal consolidations.
In contrast, the livers of the IN-infected animals (numbers 16–20) exhibited multifocal steatosis; some had a periportal distribution, while others were mainly microvesicular in pattern. In addition, four of the five animals exhibited diffuse hepatocytic necrosis; only one liver (animal 19) had centrilobular necrosis. The characteristic cytoplasmic inclusion bodies were present in all livers (Figure 1A). As observed in the IP-infected animals, moderate-to-severe splenic necrosis occurred. In addition to the variable consolidation and interstitial inflammation in the lungs, some of the IN-infected animals showed necrosis in the peribronchial lymphoid tissue. Lymph nodes from other sites (i.e., mediastinal brown fat) also showed focal necrosis, accompanied by proliferation of immunoblast-like cells, fibroblasts, and macrophages.
Immunohistochemically, no positive staining was observed in control animals. Sections of adrenals, kidneys, and hearts from the infected ground squirrels were also negative. In the liver, most of the larger inclusion bodies stained strongly positive for viral antigen (Figure 1C); however, some of the smaller inclusion bodies were negative. Depending on the severity of the histologic abnormality, this positive staining sometimes involved the surrounding cytoplasm and cytoplasmic membranes. The spleen also stained strongly positive; the intensity generally corresponded to the severity of pathologic changes (Figure 1D). In some animals, the cells lining the surface of the splenic capsule (mesothelial cells) were enlarged and were also strongly positive for viral antigen. In these animals, the positive staining appeared to extend into the superficial zones of the neighboring tissues or organs, such as fat (Figure 1E), pancreas, and adrenal gland, which otherwise were generally negative for viral antigen and lacked pathologic changes. This finding suggests that virus spread directly between adjacent sites when the boundaries (capsules) were broken. Necrotic areas in the perisplenic and periadrenal fat also stained strongly positive.
Viral antigen staining in other organs was less consistent. In the lungs, scattered interstitial cells and a few alveolar pneumocytes were positive (Figure 1F). In the kidneys, sometimes rare mononuclear leukocytes in a few glomeruli were positive. However, these latter positive monocytes probably represented cells in circulation, rather than actual virus replication in the renal tissue.
Figure 2
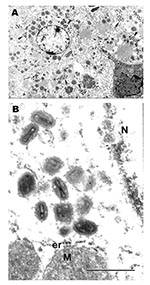
Figure 2. Ultrastructural localization of monkeypox virus in hepatocytes in the liver of a ground squirrel 5 days after infection. A) Hepatocytes contain numerous groups of virions (arrows) in their cytoplasm (bar =...
Examination of selected tissues by EM confirmed the results of immunohistochemistry. In ultrathin sections, groups of poxlike virions were readily seen within cytoplasm of infected hepatocytes (Figures 2A and B).
Results of our study indicate that the thirteen-lined ground squirrel is highly susceptible to MPXV. Experimental infection of the animals by both IP and IN routes produced a fulminant uniformly fatal disease. All of the animals were dead by the day 9. The amount and wide distribution of virus in various organs indicate that the infection was disseminated. Initially, the first recovery of MPXV from the blood of IP-infected squirrels and from the oropharynx of IN-infected animals suggested that the pathogenesis of MPXV might be different, depending on the route of infection; however, at necropsy, the amount of virus present in the respective organ systems of the two groups was similar. The histopathology observed at necropsy in the two groups was also similar, although squirrels in the IN-infected group had more hepatic steatosis and pulmonary consolidation. However, this difference may simply be a reflection of the longer incubation period and later death of the IN-infected animals.
In a recent publication, Guarner et al. (4) described the histopathologic findings in two sick prairie dogs (Cynomys spp), collected from a pet store during the 2003 monkeypox outbreak in the United States. The abnormal pathologic findings in these two animals were ulcerative lesions on the tongue and conjunctiva and in the lung (bronchioalveolar pneumonia). MPXV was recovered from the lungs of both animals, but only mild inflammation in the liver and reactive hyperplasia in the spleen were found. In another study, we experimentally infected eight prairie dogs with MPXV by the IN and IP routes. Skin or mucosal lesions developed in some of these animals; some of them survived; in general, the survival time was longer and the degree of pathology was less than that observed in the ground squirrels (unpub. data). Although both rodent species are members of the family Sciuridae, these observations suggest that MPXV infection in the thirteen-lined ground squirrel is more severe than in prairie dogs.
The fulminant disease and pathology produced in S. tridecemlineatus by MPXV are similar to the pathologic findings described in experimentally infected macaques (10), which in turn are similar to life-threatening or fatal smallpox (variola virus infection) in humans (11). This similarity suggests that ground squirrels might be an excellent small-animal model for studying the pathogenesis and treatment of severe orthopoxvirus infections in people. Concern about potential bioterrorism (12) as well as recent reports of zoonotic transmission of poxviruses (13) have renewed research interest in these viruses. S. tridecemlineatus is abundant in grassland and prairie habitats in the central United States and adjacent regions of Canada (14), so supply should be plentiful. Their adult weight (140–252 g), laboratory diet, and cage requirements are similar to those of a large hamster or small guinea pig. Thus, we feel that these animals have considerable value as a laboratory model.
Dr. Tesh is professor of pathology at the University of Texas Medical Branch in Galveston. His research interests are in the epidemiology and pathogenesis of arthropodborne and emerging zoonotic viral diseases.
Acknowledgments
We are grateful to Inger K. Damon for providing MPXV, Dora Salinas for helping prepare the manuscript, and Patrick Newman and Mengyi Ye for assisting in tissue processing and immunohistochemical staining.
This work was supported by contracts NO1-AI25489, NO1-AI30027, and U54-AI57156 from the National Institutes of Health.
References
- Di Guilio DB, Eckburg PB. Human monkeypox: an emerging zoonosis. Lancet Infect Dis. 2004;4:15–25. DOIPubMedGoogle Scholar
- Centers for Disease Control and Prevention. Update: multistate outbreak of monkeypox—Illinois, Indiana, Kansas, Missouri, Ohio and Wisconsin, 2003. MMWR Morb Mortal Wkly Rep. 2003;52:642–6.PubMedGoogle Scholar
- Reed KD, Melski JW, Graham MB, Regnery RL, Sotis MJ, Wegner MV, The detection of monkeypox in humans in the Western Hemisphere. N Engl J Med. 2004;350:342–50. DOIPubMedGoogle Scholar
- Guarner J, Johnson BJ, Paddock CD, Shieh WJ, Goldsmith CS, Reynolds MG, Monkeypox transmission and pathogenesis in prairie dogs. Emerg Infect Dis. 2004;10:426–31.PubMedGoogle Scholar
- Hutin YJF, Williams RJ, Malfait P, Pebody R, Loparev VN, Ropp SL, Outbreak of human monkeypox, Democratic Republic of Congo, 1996–1997. Emerg Infect Dis. 2001;7:434–8.PubMedGoogle Scholar
- Khodakevich L, Szczeniowski M, Manbu-ma-Disu, Jezek Z, Marennikova S, Nakano J, et al. The role of squirrels in sustaining monkeypox virus transmission. Trop Geogr Med. 1987;39:115–22.PubMedGoogle Scholar
- Jazek Z, Fenner F. Human monkeypox. Basel: Karger; 1988.
- Tesh RB, Duboise SM. Viremia and immune response with sequential phlebovirus infections. Am J Trop Med Hyg. 1987;36:662–8.PubMedGoogle Scholar
- Xiao SY, Zhang H, Guzman H, Tesh RB. Experimental yellow fever virus infection in the golden hamster (Mesocricetus auratus). 2. Pathology. J Infect Dis. 2001;183:1437–44. DOIPubMedGoogle Scholar
- Zaucha GM, Jahrling PB, Geisbert TW, Swearengen JR, Hensley L. The pathology of experimental aerosolized monkeypox virus infection in cynomolgus monkeys (Macaca fasciacularis). Lab Invest. 2001;81:1581–600.PubMedGoogle Scholar
- Councilman W, Magrath B, Brinkerhoff W. The pathological anatomy and histology of variola. J Med Res. 1904;11:12–134.PubMedGoogle Scholar
- Henderson DA. Bioterrorism as a public health threat. Emerg Infect Dis. 1998;4:488–92. DOIPubMedGoogle Scholar
- Frey SE, Belshe RB. Poxvirus zoonoses-putting pocks into context. N Engl J Med. 2004;350:324–7. DOIPubMedGoogle Scholar
- Wilson DE, Ruff S. The Smithsonian book of North American Mammals. Washington: Smithsonian Institution Press; 1999. p. 436–8.
Figures
Tables
Cite This ArticleTable of Contents – Volume 10, Number 9—September 2004
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Robert B. Tesh, Department of Pathology, University of Texas Medical Branch, 301 University Boulevard, Galveston, TX 77555-0609, USA; fax: 409-747-2429
Top