Volume 14, Number 9—September 2008
Research
Forest Fragmentation as Cause of Bacterial Transmission among Nonhuman Primates, Humans, and Livestock, Uganda
Cite This Article
Citation for Media
Abstract
We conducted a prospective study of bacterial transmission among humans, nonhuman primates (primates hereafter), and livestock in western Uganda. Humans living near forest fragments harbored Escherichia coli bacteria that were ≈75% more similar to bacteria from primates in those fragments than to bacteria from primates in nearby undisturbed forests. Genetic similarity between human/livestock and primate bacteria increased ≈3-fold as anthropogenic disturbance within forest fragments increased from moderate to high. Bacteria harbored by humans and livestock were approximately twice as similar to those of red-tailed guenons, which habitually enter human settlements to raid crops, than to bacteria of other primate species. Tending livestock, experiencing gastrointestinal symptoms, and residing near a disturbed forest fragment increased genetic similarity between a participant’s bacteria and those of nearby primates. Forest fragmentation, anthropogenic disturbance within fragments, primate ecology, and human behavior all influence bidirectional, interspecific bacterial transmission. Targeted interventions on any of these levels should reduce disease transmission and emergence.
Infectious diseases transmitted among wild nonhuman primates, humans, and domestic animals pose a serious threat to wildlife conservation, human health, and animal health (1,2). For example, outbreaks of Ebola hemorrhagic fever and anthrax have caused epidemic deaths in apes and local humans in West Africa (3,4), and human paramyxoviruses have caused repeated deaths in chimpanzees in Côte d’Ivoire (5). Emerging pathogens such as these are now regarded as important drivers of primate population declines (1,6).
Although people and domestic animals have shared habitats with nonhuman primates (primates hereafter) for centuries, the dynamics of these interactions have changed dramatically over the last several decades. The destruction of tropical forests worldwide has imperiled many primates (7). Today, most primates live in remnant forest fragments and isolated protected areas within habitat mosaics of farmland, pastures, and human settlements (8,9).
Several studies have demonstrated that fragmentation of tropical forests reduces primate biodiversity and alters primate demographics and behavior (10,11). Fragmentation also alters patterns of gastrointestinal helminthic and protozoan infection in certain species (12–14). Whether host susceptibility, transmission dynamics, or a combination of these factors drive such trends remains unclear (15). The effects of fragmentation on the dynamics of pathogen transmission between primates and other species, including humans, are largely unexplored.
The goal of this study was to assess the effects of forest fragmentation on rates and patterns of bacterial transmission among wild primates, humans, and livestock, and to examine how anthropogenic and behavioral factors affect these rates and patterns across a fragmented forest landscape. We targeted Escherichia coli, a common, genetically diverse gastrointestinal bacterium transmitted by a variety of modes, including directly and through the environment (16,17). Virulent forms of E. coli are of considerable concern as emerging zoonoses (17,18), and benign forms of the bacterium provide a useful system for understanding the transmission dynamics of a range of microbes with similar biologic and epidemiologic characteristics (19,20).
By examining genetic relationships among E. coli isolates from humans, livestock, and 3 species of primates, we inferred rates of bacterial transmission among populations of these species living in or near 3 fragments that differed in their degrees of anthropogenic disturbance. Combining bacterial genetic data with surveys of local residents allowed us to identify behavioral and demographic risk factors affecting bacterial transmission between humans and primates.
Study Site
Figure 1
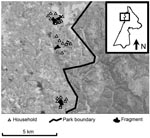
Figure 1. Map of study area within Kibale National Park, western Uganda (box) and forest fragments and households included in the study. Fragments are (from north to south) Kiko 1, Bugembe, Rurama (see...
The study took place in and near Kibale National Park, Uganda (Figure 1). Kibale is a 795-km2 park located in western Uganda near the foothills of the Rwenzori Mountains (0°13′–0°41′N, 30°19′–30°32′E), consisting primarily of moist semideciduous and evergreen forest, which is transitional between lowland rainforest and montane forest (elevation range ≈1,100–1,600 m) and interspersed with grassland, woodland, wetlands, and colonizing forest (21,22). Kibale is notable for its high species diversity and density of primates and is considered a premier primate research site in sub-Saharan Africa (23). Outside of the protected areas of Kibale exist a series of forest fragments that sustain small populations of primates (11). These fragments typically occupy nonarable wet lowlands. For this study, we focused on 3 fragments, Bugembe, Kiko 1, and Rurama, which have been studied intermittently since ≈1994 (Table 1).
Study Species
We studied the 3 species of primates typically found in fragments near Kibale: red colobus (Procolobus rufomitratus), black-and-white colobus (Colobus guereza), and red-tailed guenons (Cercopithecus ascanius). The first 2 species are folivorous and can survive even on the depauperate forest vegetation of the fragments in which they reside (11). Red-tailed guenons are omnivorous primates that favor fruit and insects in undisturbed sections of Kibale (24) but survive in nearby fragments by habitually raiding crops from adjacent farmlands (25).
Small agricultural settlements surround each fragment (Table 1, Figure 1). Persons living in these settlements are primarily subsistence farmers. Their contact with primates occurs during excursions into fragments to extract forest resources (e.g., firewood, timber) or when primates leave fragments to raid crops. Primates must also cross pastures to move among disconnected habitats within fragments, thereby coming into close contact with livestock and their feces. Livestock included in this study were cattle (Bos indicus and B. indicus x B. taurus crosses) and goats (Caprus hircus), which are ubiquitous in the region. Humans and livestock in the region use common open water sources, such as open wells and streams, which tend to be located inside or near the edges of fragments, within primate home ranges.
Sample Collection and Human Surveys
We collected fecal samples from primates (n = 93) during behavioral observations in June and July 2005 (dry season). We sampled all primate social groups from the 3 fragments as well as primates of the same species living in nearby undisturbed areas of Kibale National Park. Once a primate was observed to defecate, we recorded its species, age, sex, and individual identity (if known). We took care to sample only those portions of the fecal material that had not contacted the ground, to avoid environmental contamination. Environmental contamination from other sources (e.g., canopy vegetation) would have been unlikely, since we have been consistently unable to recover E. coli from such sources despite repeated attempts (T.L. Goldberg, unpub. data). Samples were placed in sterile tubes and transported within 6 hours to our field laboratory.
Maps and ground surveys were used to identify households within 0.5 km of each fragment; members of all of these households were invited to participate in the study in 2004. In June and July 2005, concurrent with primate sampling, members of each participating household (n = 99 persons) were given self-contained, sterile bacterial transport systems containing Cary-Blair agar (BD CultureSwab, Becton, Dickinson and Company, Franklin Lakes, NJ, USA) and were instructed in the proper method for self-administering a rectal swab. Inoculated swabs were collected and transported to our field laboratory within 24 hours of distribution. At the same time, fresh fecal samples from livestock (n = 60) owned by participating households were collected in sterile tubes and transported within 6 hours to our field laboratory.
At the time of human sample collection, a survey was administered to each participant. The survey focused on demographic information, personal health, patterns of forest use, and interactions with primates during the month before sample collection. The survey was administered in the local language by trained field assistants who were also members of the local communities; researchers were never present during survey administration to avoid response biases associated with the presence of foreigners. This study was reviewed and approved by the institutional review board and institutional animal care and use committee of the University of Illinois before data collection.
Bacterial Isolation and Characterization
Swabs and fecal samples were streaked for isolation of E. coli onto individual MacConkey agar plates and incubated at 37°C for 24 hours in our field laboratory. Up to 6 putative E. coli colonies from each sample were transferred into tubes containing 0.1 mL tryptic soy agar and stored at room temperature for up to 4 weeks. Isolates were then shipped to the University of Illinois in the United States, re-isolated, subjected to standard biochemical tests for positive identification (26), and stored in 20% glycerol at –80°C for further analysis.
Confirmed E. coli isolates were genotyped by using Rep-PCR, which targets repetitive sequences dispersed throughout bacterial chromosomes (27). This method has high power for discriminating among E. coli isolates (28,29), and it can generate accurate phylogenetic information (30). DNA extraction, PCR, and electrophoresis protocols are described in detail elsewhere (30).
Analyses
Rep-PCR genotypes were stored in the computer program BioNumerics, version 4.0 (Applied Maths, Austin, TX, USA). Relationships among isolates were inferred from Rep-PCR genotypes by using published methods that maximize the correspondence of such inferences to the standard of multilocus sequence typing (30). Population genetic analyses available in the computer program Arlequin, version 3.0 (31), were used to measure genetic differences among bacterial subpopulations. Specifically, analysis of molecular variance (AMOVA; 32) was used to apportion genetic variation among different ecologically defined bacterial subpopulations, and the common genetic distance measure, FST (33), was used to quantify short-term genetic distances among populations of bacteria from different host species and locations. Complementary phylogenetic analyses were conducted to examine relationships among individual bacterial isolates and to infer directional interspecific transmission; these were performed with BioNumerics and the computer programs PHYLIP, version 3.57c (34) and MacClade, version 4 (35), following a previously published analytical framework (36). Regression analyses were used to investigate the effects of putative demographic and behavioral risk factors on genetic similarity between bacteria from individual humans and bacteria from the primates inhabiting that person’s associated fragment (Appendix Table).
A total of 791 E. coli isolates from 252 individual persons, livestock, and primates were analyzed, representing (in the case of humans and livestock) 29 households (Table 1). Humans ranged in age from ≈2 months to 77 years and consisted of 48% male and 52% female participants. Sample sizes of primates were low in some locations, but this was inevitable, given small primate population sizes (Table 1). Human and livestock samples represented ≈50% of households surrounding each fragment.
Figure 2
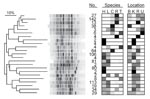
Figure 2. Dendrogram of genetic relatedness among 23 major clades of Escherichia coli from humans, domestic animals, and primates in 4 locations in and near Kibale National Park, western Uganda, derived from Rep-PCR...
Phylogenetic analysis of bacterial genotypes identified 23 major clades (Figure 2), each containing between 2 and 142 unique genotypes. Some clades contained genotypes specific to particular species or locations; others contained genotypes from multiple species and multiple locations. Of the latter type, those containing isolates from both humans and primates tended to be phylogenetically clustered (Figure 2). Phylogenetic analyses of directional interspecific transmission (36) indicated no biases in transmission for different classes of directional transmission events (e.g., human to primate, primate to human). Analyses of molecular variance (Table 2) indicated that differences among species and locations accounted for only a small proportion of total bacterial genetic diversity (7.8% and 6.8%, respectively), and that individual fragments contained most bacterial genetic diversity (85.4%).
Pairwise bacterial genetic distances between metapopulations of primates, persons, and livestock are shown in Table 3. Both humans and livestock harbored bacteria significantly more similar genetically to those of primates in fragments than to those of primates in undisturbed locations within the national park. Humans and their livestock shared very similar bacteria, as indicated by an FST of only 0.03; this genetic distance was smaller even than that between bacteria from primates in fragments and bacteria from primates in undisturbed forest (FS = 0.046), although this difference was not statistically significant.
Figure 3
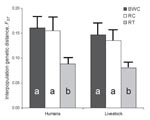
Figure 3. Interpopulation FST values between Escherica coli from humans in villages associated with 3 forest fragments near Kibale National Park, Uganda, and E. coli from livestock and primates in the same village...
Figure 4
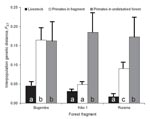
Figure 4. Interpopulation FST values between Escherichia coli from 3 species of primates in 3 forest fragments near Kibale National Park, Uganda, and E. coli from both humans and livestock living in villages...
Figure 3 shows the results of interspecies bacterial genetic distance analyses conducted separately for each fragment. Across fragments, bacteria from humans were uniformly genetically similar to bacteria from their livestock. However, genetic similarity between human and primate bacteria varied among fragments. Human–primate bacterial genetic similarity was highest in the Kiko 1 fragment, followed by Rurama, and then by Bugembe. This pattern parallels the relative degrees of anthropogenic disturbance of the fragments themselves (Kiko 1 >Rurama >Bugembe; Table 1). Species-specific analyses (Figure 4) indicated that bacteria from both humans and livestock were more similar to bacteria from red-tailed guenons than to bacteria from black-and-white colobus or red colobus.
Four variables were retained in the final regression model that examined associations between human behavioral predictors and human–primate bacterial genetic similarity (Appendix Table). Residence near a more disturbed fragment was the variable most strongly associated with increased genetic similarity between human and primate bacteria. Tending livestock, experiencing gastrointestinal symptoms, and fetching water from an open water source within the month before sampling were also associated with increased human–primate bacterial genetic similarity.
This study provides evidence that forest fragmentation increases bacterial transmission between primates and humans and their livestock. Bacteria from humans and livestock near 3 fragments were more similar genetically to bacteria from primates in those fragments than to bacteria of primates from nearby undisturbed forest locations. Moreover, the degree of disturbance of the fragments themselves paralleled the degree of genetic similarity between human and primate bacteria. Phylogenetic analyses and analyses of molecular variance further indicated that bacterial gene flow among species was high and that no directional biases in bacterial transmission were evident, findings that indicate that transmission of E. coli from primates to humans and livestock was as likely as transmission in the other direction.
Chapman et al. (15) recently showed that red colobus in forest fragments near Kibale suffer increased gastrointestinal parasitism with helminths as a result of nutritional stress and that this effect has led to a decline in population. Gillespie and Chapman (12) documented that the degree of disturbance of a fragment (measured as the density of tree stumps) was an accurate predictor of prevalence of infection of red colobus with parasitic nematodes. However, neither of these studies sampled humans or domestic animals, and neither examined transmission explicitly. The results of our study suggest that fragmentation may exert a heretofore-undocumented negative influence on the risk of primate infection by increasing pathogen transmission rates between primates and other species. Our results also show that the degree of anthropogenic disturbance within a fragment affects the rate at which bacteria are transmitted among species. Fragmentation likely leads to elevated interspecific transmission rates by increasing ecologic overlap among species.
The especially close genetic relationship between bacteria from humans and bacteria from red-tailed guenons (paralleled in livestock) probably reflects the propensity of red-tailed guenons to enter human habitats to raid crops (25). Unlike colobines, which can subsist on leaves even in highly degraded fragments, red-tailed guenons, which eat a more varied diet consisting of a high proportion of fruits, are likely obligate crop raiders in fragments. The importance of this species as a crop raider is evident from the fact that persons in our study area engage in a variety of culturally unique practices specifically designed to deter crop raiding, especially of maize, by red-tailed guenons (38).
We initially suspected that diet and digestive physiology might influence the genetic similarity of E. coli among different host species (39), but our results do not support this hypothesis. Humans in our study, who as omnivores have single stomachs, harbored E. coli virtually indistinguishable genetically from the E. coli of their cattle and goats, which are herbivores with chambered stomachs. By extension, similarities in digestive physiology between humans and red-tailed guenons would not be sufficient to account for the close genetic relationship between E. coli from these species. We infer that spatial and ecologic overlap is the primary determinant of bacterial genetic similarity among populations of hosts in our system.
Persons who tended livestock and experienced gastrointestinal symptoms during the month before sampling harbored bacteria genetically similar to those of the primates in their associated fragment, whereas persons who did not engage in these activities or experience these symptoms harbored bacteria more distantly related to those of the same primates. Tending livestock, which are often grazed in or near fragments, may bring humans into close contact with primates. Fetching water from an open water source (p = 0.07 in our regression analysis; see Appendix Table) may expose humans to water contaminated with bacteria of primate origin. We note that these variables accounted together for only 28% of variation in human–primate bacterial genetic similarity, indicating that most variation in this parameter remains unexplained.
We emphasize that the results of our risk analysis represent statistical associations and that they do not indicate direction of causality. For example, persons who experience gastrointestinal symptoms such as diarrhea may shed bacteria at high rates and thus be at increased risk of transmitting bacteria to primates; alternatively, persons who ingest microbes from primates might tend to experience gastrointestinal symptoms as a result. We also caution that our results might differ for pathogens more virulent than E. coli. For example, gastrointestinal disease would increase shedding of pathogens into the environment and affect host behavior. Our results are best interpreted as reflecting background patterns of bacterial transmission in the absence of confounders such as high virulence. Finally, we caution that assumptions inherent in our statistical analyses could affect the strength of the trends we have documented. For example, our analyses of genetic correspondence assume that parameters derived from maximum likelihood estimation are globally optimal, and our analyses of interpopulation genetic distances assume neutral molecular evolution (33).
We suspect that the patterns of bacterial genetic similarity we have documented reflect indirect transmission of microbes through the physical environment, such as through contaminated soil or water, rather than transmission by direct contact. For example, primates in the forest fragments near Kibale must come to the ground to cross open fields (often pastures) between habitat patches or to raid crops; this would increase their probability of encountering bacteria of human or livestock origin. Similarly, the location of fragments in nonarable, wet lowlands creates ideal conditions for contamination of surface water with primate feces. Unfortunately, our varied attempts to recover E. coli from water, soil, and vegetation were largely unsuccessful, perhaps because of the heat and aridity of the western Ugandan dry season (40).
Zoonotic diseases with primate origins have had global effects on human health (1). In Uganda, a high prevalence of HIV renders a significant proportion of the population immunocompromised and thus susceptible to opportunistic infections. Countries like Uganda are also undergoing rapid demographic changes and correspondingly rapid changes in land use. Our finding that a land-use change (forest fragmentation) enhances bacterial transmission between primates and an immunocompromised human population raises concerns about the potential for epidemics of zoonotic disease to originate from disturbed ecosystems such as this. Forest fragmentation may, in other words, negatively affect human public health by increasing the risks for zoonotic disease transmission from animals in forest fragments.
Forests and the primates living in them are disappearing rapidly from this region of Africa, which unfortunately typifies locations throughout the Tropics. We have already documented the extinction of 2 primate species from 1 fragment, and we predict that, without intervention, all unprotected fragments and their primates will disappear from our study area within the next 2 decades. Our results indicate that extinction of local primates may be accompanied by “spikes” in anthroponotic and zoonotic disease transmission risk, which could threaten not only the health of other primates and conservation but also human health. Mitigating these risks could entail such interventions as building closed wells, managing the grazing patterns of livestock, and encouraging the persistence of primate food trees within fragments. Understanding in greater detail how forest fragmentation and associated land-use changes affect pathogen transmission among primates, humans, and domestic animals would be critical for designing rational intervention strategies to conserve wild primates, as well as to safeguard human and animal health.
Dr Goldberg is professor of epidemiology in the Department of Pathobiological Sciences at the University of Wisconsin-Madison and director of the Kibale EcoHealth Project, Uganda. His research focuses on the ecology, epidemiology, and evolution of infectious diseases, with an emphasis on molecular epidemiology and zoonoses.
Acknowledgments
We thank the Uganda Wildlife Authority and the Uganda National Council for Science and Technology for granting us permission to conduct this research; Makerere University Biological Field Station for providing facilities and logistic support; J. Byaruhanga, P. Katurama, A. Nyamwija, J. Rusoke, and P. Omeja for providing invaluable assistance in the field; K. Inendino, K. Knuffman, and T. Tranby for providing assistance in the laboratory; and S. Paige, M. Ruiz, and W. Brown for providing assistance with spatial analyses.
This research was conducted as part of the Kibale EcoHealth Project. This material is based upon work supported by the Morris Animal Foundation under award nos. D04ZO-67 and D07Z0-024.
References
- Chapman CA, Gillespie TR, Goldberg TL. Primates and the ecology of their infectious diseases: how will anthropogenic change affect host-parasite interactions? Evol Anthropol. 2005;14:134–44. DOIGoogle Scholar
- Nunn CL, Altizer S. Infectious diseases in primates: behavior, ecology and evolution. Oxford: Oxford University Press; 2006.
- Leroy EM, Rouquet P, Formenty P, Souquiere S, Kilbourne A, Froment JM, Multiple Ebola virus transmission events and rapid decline of central African wildlife. Science. 2004;303:387–90. DOIPubMedGoogle Scholar
- Leendertz FH, Lankester F, Neel C, Drori O, Dupain J, Speede S, Anthrax joins Ebola in threatening central African great apes. Am J Primatol. 2006;68:928–33. DOIPubMedGoogle Scholar
- Kondgen S, Kuhl H, N’Goran PK, Walsh PD, Schenk S, Ernst N, Pandemic human viruses cause decline of endangered great apes. Curr Biol. 2008;18:260–4. DOIPubMedGoogle Scholar
- Leendertz FH, Pauli G, Maetz-Rensing K, Boardman W, Nunn C, Ellerbrok H, Pathogens as drivers of population declines: the importance of systematic monitoring in great apes and other threatened mammals. Biol Conserv. 2006;131:325–37. DOIGoogle Scholar
- Cowlishaw G, Dunbar R. Primate conservation biology. Chicago: University of Chicago Press; 2000.
- Chapman CA, Onderdonk DA. Forests without primates: primate/plant codependency. Am J Primatol. 1998;45:127–41. DOIPubMedGoogle Scholar
- Marsh LK, ed. Primates in fragments: ecology and conservation. New York: Kluwer Academic/Plenum Publishers; 2003.
- Estrada A, Coates-Estrada R. Tropical rain forest fragmentation and wild populations of primates at Los Tuxtlas, Mexico. Int J Primatol. 1996;17:759–83. DOIGoogle Scholar
- Onderdonk DA, Chapman CA. Coping with forest fragmentation: the primates of Kibale National Park, Uganda. Int J Primatol. 2000;21:587–611. DOIGoogle Scholar
- Gillespie TR, Chapman CA. Prediction of parasite infection dynamics in primate metapopulations based on attributes of forest fragmentation. Conserv Biol. 2006;20:441–8. DOIPubMedGoogle Scholar
- Gillespie TR, Chapman CA. Forest fragmentation, the decline of an endangered primate, and changes in host-parasite interactions relative to an unfragmented forest. Am J Primatol. 2008;70:222–30. DOIPubMedGoogle Scholar
- Salzer JS, Rwego IB, Goldberg TL, Kuhlenschmidt MS, Gillespie TR. Giardia sp. and Cryptosporidium sp. infections in primates in fragmented and undisturbed forest in western Uganda. J Parasitol. 2007;93:439–40. DOIPubMedGoogle Scholar
- Chapman CA, Wasserman MD, Gillespie TR, Speirs ML, Lawes MJ, Saj TL, Do food availability, parasitism, and stress have synergistic effects on red colobus populations living in forest fragments? Am J Phys Anthropol. 2006;131:525–34. DOIPubMedGoogle Scholar
- Donnenberg M, ed. Escherichia coli: virulence mechanisms of a versatile pathogen. San Diego: Academic Press; 2002.
- Beutin L. Emerging enterohaemorrhagic Escherichia coli, causes and effects of the rise of a human pathogen. J Vet Med B Infect Dis Vet Public Health. 2006;53:299–305. DOIPubMedGoogle Scholar
- Alekshun MN, Levy SB. Commensals upon us. Biochem Pharmacol. 2006;71:893–900. DOIPubMedGoogle Scholar
- Goldberg TL, Gillespie TR, Rwego IB, Wheeler E, Estoff EL, Chapman CA. Patterns of gastrointestinal bacterial exchange between chimpanzees and humans involved in research and tourism in western Uganda. Biol Conserv. 2007;135:511–7. DOIGoogle Scholar
- Struhsaker TT. Ecology of an African rain forest: logging in Kibale and the conflict between conservation and exploitation. Gainesville (FL): University Press of Florida; 1997.
- Chapman CA, Lambert JE. Habitat alteration and the conservation of African primates: case study of Kibale National Park, Uganda. Am J Primatol. 2000;50:169–85. DOIPubMedGoogle Scholar
- Chapman CA, Struhsaker TT, Lambert JE. Thirty years of research in Kibale National Park, Uganda, reveals a complex picture for conservation. Int J Primatol. 2005;26:539–55. DOIGoogle Scholar
- Struhsaker TT. Food habits of five monkey speices in the Kibale Forest, Uganda. In: Chivers DJ, Herbert J, editors. Recent advances in primatology. London: Academic Press; 1978. p. 225–48.
- Naughton-Treves L, Treves A, Chapman C, Wrangham R. Temporal patterns of crop-raiding by primates: linking food availability in croplands and adjacent forest. J Appl Ecol. 1998;35:596–606. DOIGoogle Scholar
- Feng P. Escherichia coli. In: Labbae RG, Garcia S, editors. Guide to foodborne pathogens. New York: John Wiley & Sons, Inc.; 2001. p. 143–62.
- Versalovic J, Koeuth T, Lupski JR. Distribution of repetitive DNA sequences in eubacteria and application to fingerprinting of bacterial genomes. Nucleic Acids Res. 1991;19:6823–31. DOIPubMedGoogle Scholar
- Johnson JR, O’Bryan TT. Improved repetitive-element PCR fingerprinting for resolving pathogenic and nonpathogenic phylogenetic groups within Escherichia coli. Clin Diagn Lab Immunol. 2000;7:265–73. DOIPubMedGoogle Scholar
- Carson CA, Shear BL, Ellersieck MR, Schnell JD. Comparison of ribotyping and repetitive extragenic palindromic-PCR for identification of fecal Escherichia coli from humans and animals. Appl Environ Microbiol. 2003;69:1836–9. DOIPubMedGoogle Scholar
- Goldberg TL, Gillespie TR, Singer RS. Optimization of analytical parameters for inferring relationships among Escherichia coli isolates from repetitive-element PCR by maximizing correspondence with multilocus sequence typing data. Appl Environ Microbiol. 2006;72:6049–52. DOIPubMedGoogle Scholar
- Excoffier L, Laval G, Schnieder S. Arlequin (version 3.0): an integrated software package for population genetics data analysis. Evol Bioinform Online. 2005;1:47–50 [cited 2008 Jun 16]. Available from http://66.102.1.104/scholar?hl=en&lr=&q=cache:pIMXAyTZ-p8J:www.la-press.com/cr_data/files/f_EBO-1-ExcoffierSc_231.pdf+Arlequin+version+3.0
- Excoffier L, Smouse PE, Quattro JM. Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics. 1992;131:479–91.PubMedGoogle Scholar
- Reynolds J, Weir BS, Cockerham CC. Estimation of the coancestry coefficient: basis for a short-term genetic distance. Genetics. 1983;105:767–79.PubMedGoogle Scholar
- Felsenstein J. PHYLIP: Phylogenetic Inference Package, version 3.57c. Seattle: Department of Genetics, University of Washington; 1995.
- Maddison DR, Maddison WP. MacClade 4: Analysis of phylogeny and character evolution, version 4.0. Sunderland (MA): Sinauer Associates; 2000.
- Goldberg TL. Application of phylogeny reconstruction and character-evolution analysis to inferring patterns of directional microbial transmission. Prev Vet Med. 2003;61:59–70. DOIPubMedGoogle Scholar
- Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4:406–25.PubMedGoogle Scholar
- Goldberg TL, Gillespie TR, Rwego IB. Health and disease in the people, primates, and domestic animals of Kibale National Park: implications for conservation. In: Wrangham R, Ross E, editors. Science and conservation in African forests: the benefits of long-term research. Cambridge: Cambridge University Press; 2008. p. 75–87.
- Hartel PG, Summer JD, Segars WI. Deer diet affects ribotype diversity of Escherichia coli for bacterial source tracking. Water Res. 2003;37:3263–8. DOIPubMedGoogle Scholar
- Rivera SC, Hazen TC, Toranzos GA. Isolation of fecal coliforms from pristine sites in a tropical rain forest. Appl Environ Microbiol. 1988;54:513–7.PubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 14, Number 9—September 2008
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Tony L. Goldberg, University of Wisconsin–Madison, Department of Pathobiological Sciences, 2015 Linden Dr, Madison, WI 53706, USA;
Top