Volume 19, Number 12—December 2013
Dispatch
Hepatitis E Virus Variant in Farmed Mink, Denmark
Cite This Article
Citation for Media
Abstract
Hepatitis E virus (HEV) is a zoonotic virus for which pigs are the primary animal reservoir. To investigate whether HEV occurs in mink in Denmark, we screened feces and tissues from domestic and wild mink. Our finding of a novel HEV variant supports previous findings of HEV variants in a variety of species.
Hepatitis E virus (HEV, family Hepeviridae) is a main cause of acute liver inflammation in humans. It is a nonenveloped RNA virus with a positive-sense genome of ≈7.2 kb. In 1997, HEV was discovered in pigs (1), and several studies have since shown that HEV is endemic in pigs and that pigs probably are a major animal reservoir. HEV traditionally had been divided into 4 primary genotypes (G1–G4). G1 and G2 have been found only in humans. G3 has been found globally in a wide range of mammals, including humans, pigs, deer, rabbits, and mongooses. G4, like G3, has an animal reservoir and has been found in humans, pigs, and wild boars (2).
Along with the human and porcine variants, avian HEV (aHEV) has been characterized. It is widespread globally and has been proposed to comprise 3 genotypes (3). Since 2010, several novel HEV variants have been described in red foxes, cutthroat trout, rats, bats, and ferrets (4–8). All new variants clearly differed from HEV G1–G4, aHEV, and each other. HEV is highly prevalent among pigs in Denmark; 92% of herds are seropositive, and ≈50% of investigated herds had pigs positive for HEV RNA (9). Because HEV is highly prevalent in swine in Denmark, animals fed offal from Danish slaughterhouses will be exposed to HEV. Production of mink fur is a major industry in Denmark, and mink are routinely fed a mixed diet, which often includes swine offal. Inappropriate heat-treated swine offal has previously been shown to be the source of swine-related influenza A virus infection in mink (10,11). Thus, we aimed to investigate whether mink in Denmark are infected with HEV G1–G4 or other HEV variants by screening fecal and tissue samples from domestic and wild mink.
Initially, we screened 85 fecal samples collected during 2006 through mid-2012 from farmed mink by nested PCR; a broad panel of HEV variants was detected (6). One sample was positive, and subsequent sequencing and phylogenetic analysis showed that this virus represented a new HEV variant. To screen more samples for this new virus, we developed a specific real-time reverse transcription PCR (RT-PCR) (Technical Appendix). The initially tested 85 fecal samples and an additional 233 fecal samples from farmed mink, together with liver and fecal samples from 89 wild mink, were tested with this new and more sensitive assay. We identified 4 positive samples, all from farmed mink. In addition, screening with an HEV real-time RT-PCR (9) specific for G1–G4 found none positive. The HEV-infected mink were all submitted for diagnostic examination; all had histories of diarrhea in the herd. Three submissions were from herds having mink enteritis virus. Lipidosis, Aleutian mink disease virus, and catarrhal enteritis also were diagnosed in the mink (online Technical Appendix Table).
The 4 samples positive for the novel HEV variant were collected during 2008–2011 from herds across Jutland, Denmark, with a minimum distance of 80 km between the herds. The 4 PCR products obtained by the nested PCR, covering a region of 261 bp of the RdRp gene, were cloned and sequenced (GenBank accession nos. KC802090, KC802091, KC802092, and KC802093). The sequences were 98%–100% identical, with only 1 nonsynonymous mutation, resulting in a neutral amino acid change from isoleucine to valine (Technical Appendix Figure). The high homology in this region is not surprisingbecause the gene encodes the RNA polymerase. We initially tried to uncover a larger fragment by primer walking, but the limited amount of material prohibited this.
Figure
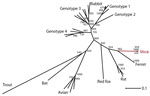
Figure. . Phylogenetic tree showing the relationship between the novel mink hepatitis E virus (HEV), other HEV variants, and the 4 known HEV genotypes based on 261 bp of the RdRp gene....
On the basis of the 261-bp fragment, we analyzed the phylogenetic relationship of this novel mink HEV variant to variants found in other animals (Figure). The mink HEV variant clustered with HEV variants found in ferrets and rats, which grouped in a separate branch that was clearly distinct from other previously described HEV variants. At nucleotide level, the mink HEV variant was ≈65% identical to the closest classical HEV genotype (G3 and G4) and 76% and 69% identical with ferret and rat HEVs, respectively. At the amino acid level, the homologies were more pronounced, showing ≈87% and ≈78% identity with ferret and rat HEV variants, respectively. The grouping of the HEV reference sequences in the analysis was identical with results of previously performed phylogenetic analysis on full-length sequences (12).
We detected a variant of HEV in 4 farmed mink from 4 geographically distinct locations in Denmark during a 3-year period, which indicates that the virus has been circulating among mink. Phylogenetic analysis showed that the virus was clearly distinct from, but closely related to, ferret and rat HEV variants recently reported from Germany and the United States (6,7,13).
It has not been possible to infect primates with rat or avian HEV variants (13,14). Thus, because of the phylogenetic resemblance of mink HEV with these nonzoonotic HEV variants, there are no indications that mink HEV can infect humans, although no human samples have been tested specifically for this virus. The zoonotic potential of HEV has been documented only in the case of G3 and G4, which were not found in mink. However, considering the relatively high HEV seroprevalence in humans, the possibility of other variants being zoonotic and cross-reacting with HEV G1–G4 in serologic assays cannot be ruled out.
The mink in this study were from herds that had mink enteritis virus, hepatic lipidosis, Aleutian mink disease virus, and catarrhal enteritis, all factors that could explain the conditions of the mink infected with HEV (15). However, it cannot be ruled out that the mink HEV variant contributed to the clinical signs of the mink HEV-positive animals. To determine whether the virus is indeed capable of inducing clinical signs in mink, the animals need to be experimentally infectedd. However, the rat and ferret HEV variants induced almost no histologic signs in rats after experimental infection, and the ferrets were described as not showing overt clinical signs (7,13). So far, only chickens infected with aHEV and humans infected with HEV G1–G4 have been described as being clinically affected by HEV infections. The possibility exists that the HEV variants recently reported in a variety of different species, including the 1 reported here, could evolve into disease-causing pathogens in animals and possibly also humans.
Dr Krog was a PhD student at the National Veterinary Institute of Denmark, and the work presented here is part of a thesis clarifying the zoonotic aspects of hepatitis E virus in Denmark. His primary research interests include HEV infection dynamics in pigs, food safety, and viral contamination of the environment.
Acknowledgments
We thank Mariann Chriél for supplying the samples from wild mink.
This project was funded by the Ministry of Food, Agriculture and Fisheries of Denmark (project no. 3304-FVFP-09-F-011).
References
- Meng X-J, Purcell RH, Halbur PG, Lehman JR, Webb DM, Tsareva TS, A novel virus in swine is closely related to the human hepatitis E virus. Proc Natl Acad Sci U S A. 1997;94:9860–5 . DOIPubMedGoogle Scholar
- Lu L, Li C, Hagedorn CH. Phylogenetic analysis of global hepatitis E virus sequences: genetic diversity, subtypes and zoonosis. Rev Med Virol. 2006;16:5–36. DOIPubMedGoogle Scholar
- Bilic I, Jaskulska B, Basic A, Morrow CJ, Hess M. Sequence analysis and comparison of avian hepatitis E viruses from Australia and Europe indicate the existence of different genotypes. J Gen Virol. 2009;90:863–73 . DOIPubMedGoogle Scholar
- Batts W, Yun S, Hedrick R, Winton J. A novel member of the family Hepeviridae from cutthroat trout (Oncorhynchus clarkii). Virus Res. 2011;158:116–23 and. DOIPubMedGoogle Scholar
- Drexler JF, Seelen A, Corman VM, Fumie Tateno A, Cottontail V, Melim Zerbinati R, Bats worldwide carry hepatitis E virus–related viruses that form a putative novel genus within the family Hepeviridae. J Virol. 2012;86:9134–47 . DOIPubMedGoogle Scholar
- Johne R, Plenge-Bönig A, Hess M, Ulrich RG, Reetz J, Schielke A. Detection of a novel hepatitis E-like virus in faeces of wild rats using a nested broad-spectrum RT-PCR. J Gen Virol. 2010;91:750–8. DOIPubMedGoogle Scholar
- Raj VS, Smits SL, Pas SD, Provacia LB, Moorman-Roest H, Osterhaus AD, Novel hepatitis E virus in ferrets, the Netherlands. Emerg Infect Dis. 2012;18:1369–70 . DOIPubMedGoogle Scholar
- Bodewes R, van der Giessen J, Haagmans BL, Osterhaus ADME, Smits SL. Identification of multiple novel viruses in feces of red foxes including a parvovirus and hepevirus. J Virol. 2013;87:7758–64. DOIPubMedGoogle Scholar
- Breum SØ, Hjulsager CK, de Deus N, Segales J, Larsen LE. Hepatitis E virus is highly prevalent in the Danish pig population. Vet Microbiol. 2010;146:144–9 . DOIPubMedGoogle Scholar
- Chriél M, Jensen TH, Hjulsager C, Larsen LE, Jørgensen PH, Harslund JL, Consequences of outbreaks of influenza A virus in farmed mink (Neovison vison) in Denmark in 2009 and 2010. In: Larsen PF, Møller SH, Clausen T, Hammer AS, Lássen TM, Nielsen VH, et al., editors. Proceedings of the Xth International Scientific Congress in Fur Animal Production. Wageningen (the Netherlands): Wageningen Academic Publishers; 2012. p. 186–9.
- Yoon K-J, Schwartz K, Sun D, Zhang J, Hildebrandt H. Naturally occurring influenza A virus subtype H1N2 infection in a Midwest United States mink (Mustela vison) ranch. J Vet Diagn Invest. 2012;24:388–91 . DOIPubMedGoogle Scholar
- Smith DB, Purdy MA, Simmonds P. Genetic variability and the classification of hepatitis E virus. J Virol. 2013;87:4161–9. DOIPubMedGoogle Scholar
- Purcell RH, Engle RE, Rood MP, Kabrane-Lazizi Y, Nguyen HT, Govindarajan S, Hepatitis E virus in rats, Los Angeles, California, USA. Emerg Infect Dis. 2011;17:2216–22 . DOIPubMedGoogle Scholar
- Huang FF, Sun ZF, Emerson SU, Purcell RH, Shivaprasad HL, Pierson FW, Determination and analysis of the complete genomic sequence of avian hepatitis E virus (avian HEV) and attempts to infect rhesus monkeys with avian HEV. J Gen Virol. 2004;85:1609–18. DOIPubMedGoogle Scholar
- Uttenthal A, Larsen S, Lund E, Bloom ME, Storgård T, Alexandersen S. Analysis of experimental mink enteritis virus infection in mink: in situ hybridization, serology, and histopathology. J Virol. 1990;64:2768–79 .PubMedGoogle Scholar
Figure
Cite This ArticleTable of Contents – Volume 19, Number 12—December 2013
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Jesper S. Krog, National Veterinary Institute, Technical University of Denmark, Bülowsvej 27, 1870 Frederiksberg, DenmarkJesper S. Krog, National Veterinary Institute, Technical University of Denmark, Bülowsvej 27, 1870 Frederiksberg, Denmark
Top