Volume 22, Number 12—December 2016
Research
Whole-Genome Characterization and Strain Comparison of VT2f-Producing Escherichia coli Causing Hemolytic Uremic Syndrome
Cite This Article
Citation for Media
Abstract
Verotoxigenic Escherichia coli infections in humans cause disease ranging from uncomplicated intestinal illnesses to bloody diarrhea and systemic sequelae, such as hemolytic uremic syndrome (HUS). Previous research indicated that pigeons may be a reservoir for a population of verotoxigenic E. coli producing the VT2f variant. We used whole-genome sequencing to characterize a set of VT2f-producing E. coli strains from human patients with diarrhea or HUS and from healthy pigeons. We describe a phage conveying the vtx2f genes and provide evidence that the strains causing milder diarrheal disease may be transmitted to humans from pigeons. The strains causing HUS could derive from VT2f phage acquisition by E. coli strains with a virulence genes asset resembling that of typical HUS-associated verotoxigenic E. coli.
Verotoxigenic Escherichia coli (VTEC) infections in humans cause a wide spectrum of clinical manifestations ranging from uncomplicated forms of intestinal illnesses to bloody diarrhea and systemic sequelae, such as hemolytic uremic syndrome (HUS) (1). The most severe forms are caused by the damage inflicted by the verocytotoxins (VTs) to the target cells in the intestinal mucosa and the renal blood vessels (1). The genes encoding the verocytotoxins (vtx) are harbored by lambdoid bacteriophages, which can be transferred to multiple bacterial hosts, generating a great diversity in the bacterial types that produce such toxins (2).
The most well-known VTEC serogroup, O157, inhabits the gastrointestinal tract of ruminants, especially cattle. However, this and other VTEC serotypes have been isolated from the feces of several other animal species, including deer, pigs, horses, cats, dogs, and wild birds (3).
During a program aimed at the control of the pigeon population in Rome, Italy during 1998, G. Dell’Omo et al. observed that this animal species was a carrier of VTEC (4). In that study, VTEC of multiple serogroups were isolated from ≈10% of the animals tested. Of 16 VTEC, 15 carried the eae gene encoding the intimin and featured genetic determinants that produced a subtype of verocytotoxin type 2 not described before, later designated VT2f (4–6). The finding of such a high prevalence of VTEC in pigeons living in Rome led to further research into these bacteria in this and other bird species worldwide. Almost all these studies succeeded in isolating VTEC, with prevalence ranging 3% to >19% in different countries and bird species; most VTEC isolated from pigeon feces and cloacal swab samples harbored the genes encoding the VT2f subtype (7–10). These findings emphasize the existence of a strict association between VTEC carrying the vtx2f genes and pigeons, which represent a reservoir for such strains.
Data on human illness attributable to VT2f-producing E. coli has been scarce until recent reports from Germany and the Netherlands described the isolation of such strains from diarrheal stool specimens from humans (11,12). Furthermore, in the Netherlands, an HUS case was recently reported to be associated with the presence of a VT2f-producing O8:H19 strain (13). We aimed to characterize at the whole-genome level 3 E. coli strains that produced the VT2f isolated from HUS and to investigate their relationships with VT2f-producing E. coli isolated from human diarrheal cases and from the pigeon reservoir.
Bacterial Strains
We investigated 22 Vt2f -producing E. coli strains. Eight previously described strains were isolated from pigeons in Italy (4); eleven strains were isolated in the Netherlands from fecal specimens from humans with diarrhea during 2008–2012 and are part of the collections held at the National Institute for Public Health and the Environment in the Netherlands (RIVM) (12). Of the 3 VT2f-producing E. coli from HUS patients, 1 was isolated in Austria in 2013 and 2 in Italy during 2013–2014. A total of 23 unrelated VTEC non-O157 strains that produced VT1 and/or VT2 subtypes other than VT2f have been used for the comparison of the profiles of virulence genes with those of the VT2f-producing isolates (Table 1).
Whole-Genome Sequencing of E. coli Strains
Sequencing of the strains isolated from fecal samples from humans with diarrhea and from pigeons was outsourced to the Central Veterinary Institute, Wageningen University (Lelystad, the Netherlands). Genome sequences were obtained by using a TruSeq protocol on an Illumina MiSeq PE300 platform (Illumina, San Diego, CA, USA). The genomes of the 3 VT2f-producing isolates from HUS patients were sequenced by using an Ion Torrent PGM (Thermo Fisher Scientific, Waltham, MA, USA) according to 400-bp protocols for library preparation through enzymatic shearing, Ion OneTouch2 emulsion PCR, enrichment, and Hi-Q sequencing kits (Thermo Fisher Scientific).
The whole-genome sequences (WGSs) of the 23 non-O157 VTEC strains are part of the European Molecular Biology Laboratory’s European Nucleotide Archive Study (http://www.ebi.ac.uk/ena; accession no. PRJEB11886). The raw reads have been subjected to quality check through FastQC and trimmed with FASTQ positional and quality trimming tool to remove the adaptors and to accept 20 as the lowest Phred value (14).
We subjected the sequences obtained with the Ion Torrent apparatus to de novo assembly by using the tool SPADES (15) and those from Illumina by using the A5 pipeline (16). The genomes have been assembled in several contigs ranging from 42 to 495 (mean 225), with N50 values (the length of the smallest contig among the set of the largest contigs that together cover at least 50% of the assembly) between 40,736 and 347,638 (mean 152,953). All the contigs were uploaded to the EMBL European Nucleotide Archive (accession no. PRJEB12203). We made annotations by using the Prokka tool (17). All the bioinformatics tools used are available on the Aries public Galaxy server (https://w3.iss.it/site/aries/).
Virulence Gene Profile Analysis and Serotyping
The presence of vtx2f and eae genes has been assessed by PCR by using primers and conditions described elsewhere (5,18). The activity of VT2f has been evaluated by Vero cell assay (VCA) as previously described (19).
We performed detection of the virulence genes cif, efa1, espABCFIJP, etpD, iha, iss, katP, lpfa, nleABC, tccP, tir, toxB, ehxA, and espP and the serotype determination in silico on the WGSs. We used blastn (available on the Aries public Galaxy server at https://w3.iss.it/site/aries/) to search databases containing the reference sequences of all the known virulence and serotype-associated genes of pathogenic E. coli (20). To perform the principal component analysis of the virulence gene profiles, we used SAS/IML studio software version 3.4 (SAS Institute, Inc., Cary, NC, USA).
We investigated plasmid profiles by using PlasmidFinder (21; https://cge.cbs.dtu.dk//services/all.php). The intimin subtyping has been performed in silico through a BLAST search (22) of the eae gene sequences from the WGS against the National Center for Biotechnology Information nucleotide repository. The intimin types of the VT2f-producing strains isolated from pigeons have been published (6,10).
rpoB Sequencing and Analysis
Amplification and sequencing of the rpoB gene were conducted to discriminate between E. coli and E. albertii species, as previously described (23). The amplicons were purified with the SureClean Plus kit (Bioline, London, UK) and sequenced using the BigDye Terminator v1.1 kit on a Genetic Analyzer 3130 (Thermo Fisher Scientific). The obtained sequences were trimmed and aligned to the reference sequences as indicated (23), using the Clustal Omega free software (http://www.ebi.ac.uk/Tools/msa/clustalo/).
Typing
We determined E. coli phylogenetic groups by using the method of Clermont et al. (24). We carried out multilocus sequence typing (MLST) of the VT2f isolates in silico according to the scheme proposed by Wirth et al. (25). We analyzed the assembled sequences by using blastn to search the MLST database downloaded from the Internet site of the MLST.UCC Mark Achtman database (http://mlst.warwick.ac.uk/mlst/dbs/Ecoli/).
Single-Nucleotide Polymorphism (SNP) Analysis
We analyzed SNPs by using the tool kSNP3 (26) available on the Galaxy project instance Aries (https://w3.iss.it/site/aries/). We set a kmer value of 23.
Characterization of the VT2f-Producing E. coli Strains
Serotyping
Of 11 VTEC strains isolated from humans with diarrhea, 5 belonged to the O63:H6 serotype. The remaining 6 isolates contained the fliCH6 (3 strains), fliCH7 (1 strain), and fliCH34 (2 strains) genes (Table 2) and belonged to serogroups O96, O113, O132, O145, and O125. For 1 isolate, the O-antigen–associated genes could not be identified (Table 2) (12).
Molecular serotyping of the 8 VT2f-producing strains isolated from pigeons showed that all the isolates had the fliCH2 and the O4, O45, O75, and O128 serogroup-associated genes. The O-antigen genes could not be identified for the isolate ED 366 (Table 2). The HUS-associated VT2f-producing E. coli strains EF453 and EF476 belonged to serotypes O80:H2 and O55:H9, respectively, while strain EF467 was O26:H11.
Virulence Gene Profiles
The E. coli strains carrying the vtx2f and isolated from pigeons have been previously reported to produce an active VT2f (6). As expected, culture supernatants from VT2f-producing strains isolated from human diarrhea and HUS induced a cytopathic effect on Vero cells morphologically compatible with that caused by verocytoxins.
All the VT2f-producing strains included in the study were positive for the eae gene (Table 2) and displayed the presence of the entire locus of enterocyte effacement (LEE) (data not shown). Most of the E. coli VT2f-producing strains isolated from diarrheal cases harbored the α-2 intimin type (8/11), followed by the β-2 (2/11) and ι (1/11) types. The VT2f-strains isolated from pigeons had been previously described to have the β-intimin (6) in most cases and, more rarely, the α-2 intimin type (10). Of 3 HUS-associated VT2f-producing strains, 2 (EF453 and EF476) carried the ξ intimin type and 1 (EF467) had the β intimin (Table 2).
All the pigeon and HUS isolates possessed the complete set of non–LEE-encoded effectors assayed (nleA, nleB and nleC) (27), whereas the isolates from human diarrhea cases displayed an unequal presence of these genes (Table 2). The efa1 gene, hallmark of the OI-122 pathogenicity island (28), was not identified in the isolates from pigeons or from human diarrheal specimens; neither were the genes ehxA, espP and katP, usually present on the large virulence plasmid of VTEC O157 and other VTEC associated with severe human disease (Table 2). However, the gene adfO, present on the OI-57 (29), was detected in all the strains investigated (Table 2).
The HUS strains EF453, EF467, and EF476 had the entire efa1 gene. Strain EF467 also had the ehxA, espP, and katP genes; the EF453 and EF476 strains had the ehxA and espP genes only (Table 2). The analysis of the plasmid profiles substantiated the finding that the 3 HUS-associated strains carried the large virulence plasmid of VTEC, revealing the presence of a sequence 100% homologous to the replicon sequence of the pO26-CRL plasmid from a VTEC O26:H- (GenBank accession no. GQ259888.1), which harbors the genes ehxA, espP, and katP.
On the basis of plasmid profiles analysis, 7 of 11 E. coli VT2f-producing strains isolated from human diarrheal feces seemed to have the replicon sequence of the plasmid pSFO (GenBank accession no. AF401292) encoding the enterohemolysin and a cluster of pap-like genes called sfp in a sorbitol-fermenting E. coli O157 (30). However, the analysis of the WGSs failed to identify the ehxA and the pap-like sequences, suggesting that the entire pSFO plasmid was not present.
Figure 1
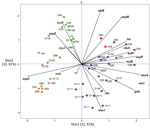
Figure 1. Principal component analysis of the virulence profiles of VT2f-producing verotoxigenic Escherichia coli (VTEC) strains isolated from fecal samples from human uncomplicated diarrheal case-patients (orange), human hemolytic uremic syndrome patients (red), and...
Principal component analysis of the virulence genes profiles showed that the HUS isolates producing VT2f clustered with the set of non-O157 VTEC isolates used for comparison, rather than with the other VT2f-producing strains (Figure 1). Conversely, the VT2f-producing strains from diarrhea and from pigeons grouped together and apart from the HUS strains (Figure 1).
Phylogenetic Analyses
rpoB Analysis
All the VT2f-producing isolates had an E. coli-related rpoB sequence (23). This finding verified that all the strains investigated were E. coli.
Typing
All VT2f-producing E. coli isolates from pigeons and the strains isolated from HUS belonged to the B1 phylogenetic group. All the strains isolated from human diarrheal feces were of phylotype B2 (Table 2).
By MLST, most of the pigeon strains investigated (5/8) belonged to sequence type (ST) 20; 1 was ST2685, and 2 were of unknown ST (Table 2), mainly because of the absence of a recognizable adk gene sequence. The 5 O63:H6, the 1 O125:H6, and the 1 ONT:H6 VTEC strains from diarrheal fecal specimens belonged to ST583; of the remaining 4 strains, 1 each was of sequence types ST28, ST121, ST582, and ST722 (Table 2).
Of 3 HUS-associated VT2f-producing E. coli, 2 (EF453 and EF476) belonged to ST301; strain EF467 was of ST21 (Table 2). All of the STs belonged to different clonal complexes or to any clonal complex, indicating that they were not related each other (data not shown).
SNP Analysis
Figure 2
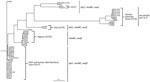
Figure 2. Parsimony tree obtained from the core-genome single-nucleotide polymorphism analysis of VT2f-producing verotoxigenic Escherichia coli (VTEC) strains from this study compared with VTEC belonging to the most relevant serogroups with publicly available...
A parsimony tree representing the core-genome SNPs analysis (Figure 2) shows that VT2f-producing strains from pigeons, human diarrheal feces, and HUS cases cluster apart from each other and from other VTEC strains used for comparison. The HUS-associated EF467 strain clusters together with the group of VTEC non-O157 from human disease, in agreement with the principal component analysis (Figures 1, 2).
Identification of a Bacteriophage Containing the vtx2f Genes
The contigs containing the vtx2f genes in the different strains ranged 2,500–68,480 bp in size. Upon annotation, they showed the presence of phage-associated genes in the proximity of vtx2f, including those encoding the antitermination protein Q, the lysis protein S, a phage terminase, an integrase, and tail-assembly proteins.
We used WGS of HUS strain EF467 to assemble a partial VT2f phage sequence of 38,594 bp (Technical Appendix). Analysis of the construct highlighted the absence of most of the genes normally involved in the regulation of the switch between the lysogenic state and the lytic cycle of lambdoid phages, such as cro, cI, cII, cIII, and N. We confirmed these findings by mapping the raw reads of the WGS of the strain EF467 against the VT2 reference bacteriophage BP933W (GenBank accession no. AF125520) (not shown).
We confirmed the genomic structure of VT2f phage obtained in silico by 4 PCRs performed on the total DNA extracted from strain EF467 by using primer pairs designed on the construct’s map and by using a restriction fragment length polymorphism analysis on the obtained PCR fragments (Table 3; Technical Appendix). The partial VT2f phage sequence was deposited into the EMBL database (accession no. LN997803).
The portion of the VT2f phage spanning the xerC gene and the tRNA-Gly and tRNA-Thr loci (Technical Appendix) was found in the draft genomes of all the VT2f-producing strains. The region downstream to the tRNA loci was also detected, but with different degrees of variation among the WGSs; for example, the presence of an additional DNA stretch of ≈24 kb in the strain BCW5746 (Technical Appendix).
Figure 3
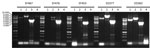
Figure 3. Long PCR analysis of the VT2f phage of verotoxigenic Escherichia coli (VTEC) strains isolated from fecal samples from humans with hemolytic uremic syndrome (EF467, EF476, EF453) and from pigeon feces (ED363,...
The presence of a similar phage structure was confirmed by using a long PCR approach in the other 2 VT2f-producing E. coli from HUS cases and in 2 strains from pigeons (strains ED377 and ED363). All the strains showed the expected amplicons with PCR 2 and 3 together with the expected product of PCR 1 in 2 pigeon isolates; all the isolates tested did not yield any amplicon with PCR 4 (Figure 3). A comparative analysis of the xerC sequences from all the VT2f phage constructs returned a high degree of variation in its sequence, explaining the observed absence of the PCR4-specific amplicon (data not shown). Finally, a BLAST search by using this VT2f phage construct from strain EF467 returned only partial similarity with phages identified in different Enterobacteriaceae but did not retrieve similar structures.
VTEC producing the VT2f subtype have long been considered a minor public health problem because of their rare association with human infections (31–34). Recently, however, an increasing number of reports of human diseases caused by infection with these E. coli strains have populated the literature (11,12,32,35). The diversity of vtx2f gene sequences compared with other vtx2 subtype genes may have played a role in underestimating the global burden of such infections. PCR primers mostly used for the detection of vtx2 genes in clinical specimens and the vehicles of infection have been proven to be unable to amplify the vtx2f gene (36). In addition, the recent description of another eae-positive Escherichia species often isolated from birds, and sometimes carrying vtx2f genes, E. albertii, added a further element of confusion. E. albertii has been associated both with gastroenteritis in humans and with healthy and diseased birds (37,38), but this species is difficult to distinguish from E. coli when using the usual biochemical or molecular assays.
Most human infections with VTEC producing VT2f have been reported as uncomplicated diarrheal cases (11,12), which may also have accounted for the underestimation of these infections. Because such cases are not actively surveyed in many countries, these infections may have been overlooked. The recent description of an HUS case associated with a VT2f-producing E. coli (13) changed the perspective on VT2f-producing E. coli and the associated disease, making it necessary to update the current paradigm of HUS-associated VTEC.
We provide evidence that the VT2f-producing E. coli isolated from HUS cases display the complete set of virulence genes described in the typical HUS-associated VTEC (Table 2; Figure 1) (28). All VT2f strains from HUS that we examined were positive for pathogenicity island OI-122 (28) and the large virulence plasmid first described in VTEC O157 (Table 2) (39); the strains from pigeons or from humans with uncomplicated diarrhea did not have these virulence-associated mobile genetic elements (Table 2) (10–12,32,40).
Our study also showed that the LEE was complete in all the genomes investigated, but a complete set of nleABC genes was found only in strains from pigeons and from humans with HUS (Table 2), indicating that the VT2f-producing isolates investigated belonged to 3 distinct main virulotypes or subpopulations (Table 2). The intimin subtyping supported this observation. Of 11 diarrheal isolates, 8 had the α-2 gene; all the pigeon isolates had a β intimin coding gene. Finally, 2 of the 3 strains from HUS showed the presence of a gene encoding the ξ intimin (Table 2). Furthermore, the analysis of core genome SNPs confirmed the existence of different subpopulations of VT2f-producing E. coli (Figure 2). The analysis of the virulence genes suggests that different populations of VT2f-producing E. coli exist and have different potential to cause human disease on the basis of the virulotype to which they belong.
VT2f-producing E. coli strains isolated from uncomplicated human cases of diarrhea have been reported in the literature as being ST20 (11), which is the same sequence type we identified in most pigeon isolates; this ST was also described in VT2f-producing E. coli isolated from pigeons in Japan (40). The same study also described an animal isolate of ST722, which was found in 1 strain isolated from human diarrheal feces in our study (Table 2). Similarly, the serotypes in some cases appeared to overlap isolates from pigeons and human cases of diarrhea, such as the serotype O128:H2 that we found in 1 pigeon isolate that was also reported in isolates from human cases of diarrhea in Germany (11).
Altogether, these observations indicate that the VT2f-producing E. coli causing diarrhea in humans could be a subpopulation of those inhabiting the pigeon reservoir. Alternately, information on the serotypes, ST, and principal component analysis of virulence genes profiles supports the hypothesis that the HUS VT2f-producing strains are more similar to the non-O157 VTEC often isolated from samples from humans with severe disease (Figure 1) than to the other VT2f-producing E. coli from humans with diarrhea or from asymptomatic pigeons. This hypothesis suggests that the HUS VT2f-producing strains represent a distinct population of VTEC; whether they are part of the pigeon intestinal flora or arise from an acquisition of the vtx2-phage is difficult to ascertain.
The phylogeny of VTEC of different serogroups, investigated by core SNP analysis, showed that the different VT2f-producing E. coli cluster into different subpopulations that include strain EF467 grouping together with non-O157 VTEC strains from humans with disease (Figure 2). However, the results from SNP analysis for VTEC of multiple serogroups should be carefully evaluated; the population structure of VTEC belonging to serogroups other than O157 and O26 has not been completely investigated yet.
At the first characterization of the vtx2f genes, it was proposed that they were, similar to other VT-coding genes, located on bacteriophages (5). Our study confirms this hypothesis and shows that such a phage apparently does not have similar counterparts in the VT-phage genomes reported in the National Center for Biotechnology Information nucleotide repository (http://www.ncbi.nlm.nih.gov/). In addition, we observed that VT2f phage was very similar in all the VT2f-producing E. coli investigated (Figure 3; Technical Appendix), suggesting that the vtx2f genes are present in phages sharing a common ancestor that is different from other phages with the other vtx1/vtx2 subtypes.
In conclusion, we provide evidence that human infections with VT2f-producing E. coli are zoonotic diseases transmitted from pigeons. Such an animal reservoir may either directly disseminate VTEC strains causing diarrhea or indirectly release VT2f phages in the environment, which can in turn lysogenize E. coli strains that contain accessory virulence determinants and confer them the ability to cause HUS. The isolation of VT2f-producing E. coli with a virulence gene profile related to the other HUS-associated VTEC suggests that the severity of the symptoms induced by infection may depend more on the ability to achieve a proficient colonization of the host gut mucosa rather than on the subtype of the produced toxin.
Dr. Grande is a researcher in the field of molecular microbiology. Most of her research activities have been at the European Union Reference Laboratory for E. coli, in the unit of Foodborne Zoonoses of the Italian National Institute of Health in Rome, Italy. Her research interests include the investigation and characterization of mobile genetic elements encoding virulence determinants in pathogenic E. coli.
References
- Tozzoli R, Scheutz F. Diarrheagenic Escherichia coli infections in umans. In: Morabito S, editor. Pathogenic Escherichia coli: molecular and cellular microbiology. Poole (UK): Caister Academic Press; 2014. p. 1–18.
- Tozzoli R, Grande L, Michelacci V, Ranieri P, Maugliani A, Caprioli A, Shiga toxin-converting phages and the emergence of new pathogenic Escherichia coli: a world in motion. Front Cell Infect Microbiol. 2014;4:80.DOIPubMedGoogle Scholar
- Caprioli A, Morabito S, Brugère H, Oswald E. Enterohaemorrhagic Escherichia coli: emerging issues on virulence and modes of transmission. Vet Res. 2005;36:289–311.DOIPubMedGoogle Scholar
- Dell’Omo G, Morabito S, Quondam R, Agrimi U, Ciuchini F, Macrì A, Feral pigeons as a source of verocytotoxin-producing Escherichia coli. Vet Rec. 1998;142:309–10.DOIPubMedGoogle Scholar
- Schmidt H, Scheef J, Morabito S, Caprioli A, Wieler LH, Karch H. A new Shiga toxin 2 variant (Stx2f) from Escherichia coli isolated from pigeons. Appl Environ Microbiol. 2000;66:1205–8.DOIPubMedGoogle Scholar
- Morabito S, Dell’Omo G, Agrimi U, Schmidt H, Karch H, Cheasty T, Detection and characterization of Shiga toxin-producing Escherichia coli in feral pigeons. Vet Microbiol. 2001;82:275–83.DOIPubMedGoogle Scholar
- Nielsen EM, Skov MN, Madsen JJ, Lodal J, Jespersen JB, Baggesen DL. Verocytotoxin-producing Escherichia coli in wild birds and rodents in close proximity to farms. Appl Environ Microbiol. 2004;70:6944–7.DOIPubMedGoogle Scholar
- Farooq S, Hussain I, Mir MA, Bhat MA, Wani SA. Isolation of atypical enteropathogenic Escherichia coli and Shiga toxin 1 and 2f-producing Escherichia coli from avian species in India. Lett Appl Microbiol. 2009;48:692–7.PubMedGoogle Scholar
- Kobayashi H, Kanazaki M, Hata E, Kubo M. Prevalence and characteristics of eae- and stx-positive strains of Escherichia coli from wild birds in the immediate environment of Tokyo Bay. Appl Environ Microbiol. 2009;75:292–5.DOIPubMedGoogle Scholar
- Askari Badouei M, Zahraei Salehi T, Koochakzadeh A, Kalantari A, Tabatabaei S. Molecular characterization, genetic diversity and antibacterial susceptibility of Escherichia coli encoding Shiga toxin 2f in domestic pigeons. Lett Appl Microbiol. 2014;59:370–6.DOIPubMedGoogle Scholar
- Prager R, Fruth A, Siewert U, Strutz U, Tschäpe H. Escherichia coli encoding Shiga toxin 2f as an emerging human pathogen. Int J Med Microbiol. 2009;299:343–53.DOIPubMedGoogle Scholar
- Friesema I, van der Zwaluw K, Schuurman T, Kooistra-Smid M, Franz E, van Duynhoven Y, Emergence of Escherichia coli encoding Shiga toxin 2f in human Shiga toxin-producing E. coli (STEC) infections in the Netherlands, January 2008 to December 2011. Euro Surveill. 2014;19:26–32.DOIPubMedGoogle Scholar
- Friesema IH, Keijzer-Veen MG, Koppejan M, Schipper HS, van Griethuysen AJ, Heck ME, Hemolytic uremic syndrome associated with Escherichia coli O8:H19 and Shiga toxin 2f gene. Emerg Infect Dis. 2015;21:168–9.DOIPubMedGoogle Scholar
- Cuccuru G, Orsini M, Pinna A, Sbardellati A, Soranzo N, Travaglione A, Orione, a web-based framework for NGS analysis in microbiology. Bioinformatics. 2014;30:1928–9.DOIPubMedGoogle Scholar
- Bankevich A, Nurk S, Antipov D, Gurevich AA, Dvorkin M, Kulikov AS, SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 2012;19:455–77.DOIPubMedGoogle Scholar
- Tritt A, Eisen JA, Facciotti MT, Darling AE. An integrated pipeline for de novo assembly of microbial genomes. PLoS One. 2012;7:e42304.DOIPubMedGoogle Scholar
- Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics. 2014;30:2068–9.DOIPubMedGoogle Scholar
- Paton AW, Paton JC. Detection and characterization of Shiga toxigenic Escherichia coli by using multiplex PCR assays for stx1, stx2, eaeA, enterohemorrhagic E. coli hlyA, rfbO111, and rfbO157. J Clin Microbiol. 1998;36:598–602.PubMedGoogle Scholar
- Caprioli A, Nigrelli A, Gatti R, Zavanella M. Isolation of verotoxin-producing Escherichia coli from slaughtered pigs. Eur J Clin Microbiol Infect Dis. 1993;12:227–8.DOIPubMedGoogle Scholar
- Joensen KG, Tetzschner AM, Iguchi A, Aarestrup FM, Scheutz F. Rapid and easy in silico serotyping of Escherichia coli isolates by use of whole-genome sequencing data. J Clin Microbiol. 2015;53:2410–26.DOIPubMedGoogle Scholar
- Carattoli A, Zankari E, García-Fernández A, Voldby Larsen M, Lund O, Villa L, In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob Agents Chemother. 2014;58:3895–903.DOIPubMedGoogle Scholar
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–10.DOIPubMedGoogle Scholar
- Lindsey RL, Fedorka-Cray PJ, Abley M, Turpin JB, Meinersmann RJ. Evaluating the occurrence of Escherichia albertii in chicken carcass rinses by PCR, Vitek analysis, and sequencing of the rpoB gene. Appl Environ Microbiol. 2015;81:1727–34.DOIPubMedGoogle Scholar
- Clermont O, Christenson JK, Denamur E, Gordon DM. The Clermont Escherichia coli phylo-typing method revisited: improvement of specificity and detection of new phylo-groups. Environ Microbiol Rep. 2013;5:58–65.DOIPubMedGoogle Scholar
- Wirth T, Falush D, Lan R, Colles F, Mensa P, Wieler LH, Sex and virulence in Escherichia coli: an evolutionary perspective. Mol Microbiol. 2006;60:1136–51.DOIPubMedGoogle Scholar
- Gardner SN, Slezak T, Hall BG. kSNP3.0: SNP detection and phylogenetic analysis of genomes without genome alignment or reference genome. Bioinformatics. 2015;31:2877–8.DOIPubMedGoogle Scholar
- Dean P, Kenny B. The effector repertoire of enteropathogenic E. coli: ganging up on the host cell. Curr Opin Microbiol. 2009;12:101–9.DOIPubMedGoogle Scholar
- Karmali MA, Mascarenhas M, Shen S, Ziebell K, Johnson S, Reid-Smith R, Association of genomic O island 122 of Escherichia coli EDL 933 with verocytotoxin-producing Escherichia coli seropathotypes that are linked to epidemic and/or serious disease. J Clin Microbiol. 2003;41:4930–40.DOIPubMedGoogle Scholar
- Imamovic L, Tozzoli R, Michelacci V, Minelli F, Marziano ML, Caprioli A, OI-57, a genomic island of Escherichia coli O157, is present in other seropathotypes of Shiga toxin-producing E. coli associated with severe human disease. Infect Immun. 2010;78:4697–704.DOIPubMedGoogle Scholar
- Brunder W, Karch H, Schmidt H. Complete sequence of the large virulence plasmid pSFO157 of the sorbitol-fermenting enterohemorrhagic Escherichia coli O157:H- strain 3072/96. Int J Med Microbiol. 2006;296:467–74.DOIPubMedGoogle Scholar
- Friedrich AW, Bielaszewska M, Zhang WL, Pulz M, Kuczius T, Ammon A, Escherichia coli harboring Shiga toxin 2 gene variants: frequency and association with clinical symptoms. J Infect Dis. 2002;185:74–84.DOIPubMedGoogle Scholar
- Jenkins C, Willshaw GA, Evans J, Cheasty T, Chart H, Shaw DJ, Subtyping of virulence genes in verocytotoxin-producing Escherichia coli (VTEC) other than serogroup O157 associated with disease in the United Kingdom. J Med Microbiol. 2003;52:941–7.DOIPubMedGoogle Scholar
- Seto K, Taguchi M, Kobayashi K, Kozaki S. Biochemical and molecular characterization of minor serogroups of Shiga toxin-producing Escherichia coli isolated from humans in Osaka prefecture. J Vet Med Sci. 2007;69:1215–22.DOIPubMedGoogle Scholar
- van Duynhoven YT, Friesema IH, Schuurman T, Roovers A, van Zwet AA, Sabbe LJ, Prevalence, characterisation and clinical profiles of Shiga toxin-producing Escherichia coli in The Netherlands. Clin Microbiol Infect. 2008;14:437–45.DOIPubMedGoogle Scholar
- Buvens G, De Rauw K, Roisin S, Vanfraechem G, Denis O, Jacobs F, Verocytotoxin-producing Escherichia coli O128ab:H2 bacteremia in a 27-year-old male with hemolytic-uremic syndrome. J Clin Microbiol. 2013;51:1633–5.DOIPubMedGoogle Scholar
- Feng PC, Jinneman K, Scheutz F, Monday SR. Specificity of PCR and serological assays in the detection of Escherichia coli Shiga toxin subtypes. Appl Environ Microbiol. 2011;77:6699–702.DOIPubMedGoogle Scholar
- Oh JY, Kang MS, Hwang HT, An BK, Kwon JH, Kwon YK. Epidemiological investigation of eaeA-positive Escherichia coli and Escherichia albertii strains isolated from healthy wild birds. J Microbiol. 2011;49:747–52.DOIPubMedGoogle Scholar
- Ooka T, Tokuoka E, Furukawa M, Nagamura T, Ogura Y, Arisawa K, Human gastroenteritis outbreak associated with Escherichia albertii, Japan. Emerg Infect Dis. 2013;19:144–6.DOIPubMedGoogle Scholar
- Karch H, Heesemann J, Laufs R, O’Brien AD, Tacket CO, Levine MM. A plasmid of enterohemorrhagic Escherichia coli O157:H7 is required for expression of a new fimbrial antigen and for adhesion to epithelial cells. Infect Immun. 1987;55:455–61.PubMedGoogle Scholar
- Murakami K, Etoh Y, Ichihara S, Maeda E, Takenaka S, Horikawa K, Isolation and characteristics of Shiga toxin 2f-producing Escherichia coli among pigeons in Kyushu, Japan. PLoS One. 2014;9:e86076.DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 22, Number 12—December 2016
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Laura Grande, European Union Reference Laboratory for E. coli, Veterinary Public Health and Food Safety Department, Istituto Superiore di Sanità, Viale Regina Elena 299, 00161 Rome, Italy
Top