Volume 32, Number 5—May 2026
Dispatch
One Health Investigation into Fatal Encephalitis Caused by Pigeon Paramyxovirus Type 1, France
Cite This Article
Citation for Media
Abstract
Pigeon paramyxovirus type 1 (PPMV-1) is a genotype of avian paramyxovirus type 1 that uses species of the family Columbidae as reservoir species. We report fatal PPMV-1 encephalitis in a human without immunosuppression or travel history outside metropolitan France. Postmortem analyses revealed PPMV-1 in tissues, underscoring that physicians should consider this potential diagnosis.
Paramyxoviridae is a family of enveloped, negative-sense single-stranded RNA viruses that includes many human pathogens. Paramyxoviruses have a broad host range and high risk for spillover events to humans (1). Newcastle disease virus, classified as Avian orthoavulavirus (AOAV-1; formerly avian paramyxovirus type 1 [APMV-1]), is divided into class I and II, which are further subdivided into genotypes and subgenotypes (2–4). Pigeon paramyxovirus type 1 (PPMV-1) refers to class II genotype VI strains that circulate in pigeons and doves (5,6). The first well-documented human PPMV-1 infections were reported in the late 1990–2000s as fatal pneumonia in immunosuppressed transplant recipients (7–9). During the past few years, 2 fatal cases of PPMV-1–associated encephalitis were reported in immunodeficient patients (10,11). We report an autochthonous case of fatal encephalitis caused by PPMV-1 in France.
A 69-year-old man was admitted to an emergency department after repeated falls at home. A week earlier, he attended a shamanism workshop in the Ardèche forest (France). Four days before hospitalization, marked asthenia, diarrhea, diplopia, and dizziness developed in the patient, resulting in falls without loss of consciousness. His family reported a cough for the past year and fatigue and weight loss for several months. The patient had never traveled outside metropolitan (mainland) France.
Figure 1
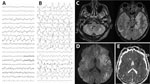
Figure 1. Electroencephalogram (EEG) and brain magnetic resonance imaging (MRI) testing of a patient with encephalitis caused by pigeon paramyxovirus type 1, France. A) The first EEG (day 5 after hospitalization) revealed...
At admission, the patient was confused and unable to stand or walk and had a fever of 39°C, right-ear hearing loss, right peripheral facial paralysis, left upper limb ataxia, and multidirectional nystagmus. Deep tendon and plantar reflexes were unremarkable. Routine laboratory tests revealed mild inflammation (hyperleukocytosis at 15 G/L [reference range 4–10 G/L]) and C-reactive protein of 25 mg/L (reference range <5 mg/L). Two cerebrospinal fluid (CSF) analyses revealed no pleocytosis, negative viral and bacterial PCR testing, and negative cultures. We noted isolated hyperproteinorachia of 0.72 g/L (reference range 0.15–0.45 g/L). An electroencephalogram revealed nonspecific encephalopathy (Figure 1, panel A). Brain magnetic resonance imaging revealed no hemorrhage or recent ischemia but showed mild white matter hyperintense lesions at the junction of the mesencephalon and pons on T2 or fluid-attenuated inversion recovery.
Five days after admission, the patient was transferred to the intensive care unit for coma, respiratory failure, and aspiration pneumonia. His neurologic condition deteriorated, and dysarthria, diplopia, deafness, ataxia, swallowing disorders, and quadriparesis developed. He was promptly intubated and mechanically ventilated. Repeat electroencephalogram (Figure 1, panel B) and brain magnetic resonance imaging (Figure 1, panels C–E) confirmed severe encephalitis. A third CSF analysis confirmed hyperproteinorachia of 1.5 g/L and no pleocytosis. Routine laboratory test results were unremarkable, including investigations for rare etiologies of encephalitis (Appendix). Electroneuromyography revealed severe motor and sensory axonal neuropathy of all 4 limbs. A full-body computed tomography scan revealed enlarged centimetric mediastinal and hilar lymphadenopathies and widespread colitis from the sigmoid to the cecum. Despite supportive care and intensive medical management (Appendix), the patient died 26 days after admission from progressively worsening, life-threatening encephalitis and polyradiculoneuropathy of unknown etiology.
To investigate the origin of the encephalitis, we performed metagenomic next-generation sequencing on postmortem midbrain and cervical spinal cord tissues obtained at autopsy, generating 82.5 million midbrain reads and 67.3 million cervical spinal cord reads. We assembled the complete PPMV-1/Human/France/2023 genome from 3,245 reads (0.48%, sequencing depth 28×) from the midbrain and 3,790 reads (0.68%, sequencing depth 34×) from the cervical spinal cord samples. We did not detect any additional pathogens.
Figure 2

Figure 2. Phylogenetic analysis of fusion gene sequences of PPMV-1from a human case of fatal PPMV-1encephalitis in France and representative APMV-1 strains. A) We conducted phylogenetic analysis on the basis of the...
Phylogenetic analyses revealed that PPMV-1/Human/France/2023 belongs to APMV-1 class II, genotype VI, sub-genotype 2.1.1.2.2. The predicted fusion protein contained the polybasic cleavage motif 112RRQKRF117 associated with virulence in birds (10,12). PPMV-1/Human/France/2023 was related to PPMV-1 strains from China, Egypt, Belgium, and Ukraine but also APMV-1/pigeon/France/172784/2017 from France (Haute-Garonne department). The sequence from France was obtained through a surveillance program in wild birds: the original sample was collected from a wild pigeon in November 2017, ≈250 km away from Ardèche (Appendix Table 1). PPMV-1/Human/France/2023 is more closely related to Pi/SH/CH/041002/2011 (GenBank accession no. PP297102.1), which was discovered in pigeons from China in 2011 (Figure 2; Appendix Table 2).
Figure 3
![PPMV-1 dissemination in the central and peripheral nervous systems and in axillary lymph nodes in a patient with fatal PPMV-1 encephalitis in France. A) Schematic representation of PPMV-1 viral load in 30 postmortem tissue samples; complete list of all samples is provided (Appendix Table 3, https://wwwnc.cdc.gov/EID/article/32/5/25-1576-App1.pdf). Viral loads were estimated at 5.2 × 105−2.6 × 106 genome copies/gram of tissue in the midbrain and 1.1−5.5 × 105 genome copies/gram of tissue in the cervical spinal cord. B) PPMV-1 viral burden in different anatomic compartments. Samples were classified into 6 groups: group 1, forebrain (including telencephalon and diencephalon); group 2, midbrain; group 3, hindbrain (including metencephalon and myelencephalon); group 4, spinal cord and spinal ganglion; group 5, lymph nodes; group 6, peripheral nervous system and peripheral samples. For the 17 forebrain samples (group 1), the PPMV-1 Ct values were the lowest (median 19.76 [IQR 16.90–25.62]), reflecting the highest viral burden. A Ct value >30 was observed in only 2 forebrain samples, the choroid plexus (Ct = 32.2) and the pituitary gland (Ct = 33.85). The 3 midbrain samples (group 2) and the 2 hindbrain samples (group 3) had higher Ct values: medians 24.05 (IQR 23.77–25.63) and 24.17 (IQR 21.33–27.00). Ct value further increased in the subsequent groups: medians 30.91 (IQR 27.49–34.39) in the 4 spinal cord and 2 spinal ganglion samples (group 4) and 36.49 (IQR 36.06–36.92) in the 2 axillary lymph node samples (group 5), indicating a progressive decrease in viral burden. The last group, consisting of 17 samples (group 6) including peripheral nervous system samples and 5 peripheral samples were all negative. We conducted a nonparametric Kruskal-Wallis test that indicated differences among groups (p<0.0001). Posthoc tests revealed an increasing trend in PPMV-1 Ct value from the brain to the peripheral samples. Pairwise Wilcoxon tests (2-tailed) for differences in means between groups adjusted with Bonferroni correction are displayed. Nonsignificant p values (>0.05) are not shown. Horizontal line within boxes indicate medians, box tops and bottoms represent IQRs, and whiskers indicate minimum and maximum values. Ct, cycle threshold; IQR, interquartile range; PPMV-1, pigeon paramyxovirus type 1.](/eid/images/25-1576-F3-tn.jpg)
Figure 3. PPMV-1 dissemination in the central and peripheral nervous systems and in axillary lymph nodes in a patient with fatal PPMV-1 encephalitis in France. A) Schematic representation of PPMV-1 viral load...
To investigate the dissemination of PPMV-1 in the patient, we conducted semiquantitative real-time PCR on all clinical samples (Appendix Table 3). PPMV-1 was detected in the CNS, thoracic spinal ganglia, and axillary lymph nodes. We identified the highest viral burdens in the brain, specifically localized to the forebrain. Posthoc tests revealed an increasing trend in PPMV-1 cycle threshold values from the brain to the peripheral samples (Figure 3).
Figure 4
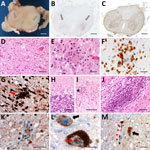
Figure 4. Histologic examination of the nervous tissue in fatal human encephalitis case caused by pigeon paramyxovirus type 1, France. A) Brown discoloration of the anterior horns (gray arrows) of the spinal...
Postmortem examination of the CNS revealed brain edema, friable midbrain tissue, and gray matter discoloration of the spinal cord (Appendix). Histopathologic testing (Appendix Table 4) revealed a diffuse inflammatory infiltrate of activated macrophages, microglia, and CD3+ and CD8+ T lymphocytes, with severe neuronal loss in the spinal cord, brainstem, and cerebellum. Neuronophagia and microglial nodules were frequent. We did not observe cytopathic effect. The choroid plexus and ependymal cells appeared unremarkable. We found perivascular inflammatory infiltrates in peripheral nerves (Figure 4). Newcastle disease virus immunostaining was positive in the spinal cord, midbrain, cerebellum, hypothalamus, and lymph nodes (Figure 4).
We report a case of fatal encephalitis associated with PPMV-1 in France that belonged to virulent class II, genotype VI, subgenotype 2.1.1.2.2, with polybasic F cleavage motif RRQKRF, in a patient with no known immunosuppression. That motif is associated with high intracerebral pathogenicity in birds (13) and was reported in another fatal encephalitis case (10). PPMV-1 belongs to the velogenic pathotype of AOAV-1 (Appendix).
The patient had never traveled outside France, indicating autochthonous infection. Although the source of infection is undetermined, the temporal association with a shamanism workshop in the Ardèche forest suggests environmental exposure to avian pathogens, potentially through contact with avian feces. PPMV-1 is shed orally and cloacally, remains stable in pigeon feces, and can spread by windborne dust, extending risk beyond localized environments (8).
Two previously reported human APMV-1 encephalitis cases occurred in severely immunosuppressed patients, suggesting that immune status might modulate APMV-1 neuropathogenicity (10,11). However, the patient we report had no known underlying disease or immunosuppressive treatment. His family reported a deterioration in his general condition, including severe weight loss, in the year before death, without medical evaluation. No laboratory markers of immunosuppression were found. We quantified blood torque teno virus loads to further assess immunosuppression, but results were inconclusive regarding immune status at the time of PPMV-1 infection (14). Detecting unusual pathogens in seemingly immunocompetent patients should prompt immune status investigation.
Real-time PCR of postmortem and peripheral samples exhibited strong neurotropism of PPMV-1 without meningitis or systemic infection at the time of neurologic manifestations. PPMV-1 was not detected in the CSF by PCR or metagenomics. PCR of bronchoalveolar lavage was negative 10 days after symptom onset, suggesting weak and transient respiratory replication despite the respiratory tract being the usual entry route. Negative blood PCR results on days 16 and 24 might reflect a brief early viremia.
The patient’s clinical manifestations included severe polyradiculoneuropathy with inflammatory infiltrates in the spinal cord and peripheral nerves, which were not described previously (10,11). The patient also exhibited enlarged lymphadenopathies, and PPMV-1 was detected at low burden in the right and left axillary lymph nodes; immunostaining confirmed viral tropism in those nodes. Lymph node aspiration could provide a minimally invasive alternative to brain biopsy for diagnosing PPMV-1 encephalitis.
In summary, in the case we report and 2 other reported encephalitis cases, class II APMV-1 with a virulence-associated F-protein cleavage site was identified. APMV-1 infection might be underdiagnosed and should be considered in neurologic disease of unknown etiology, particularly among patients who report exposure to Columbidae fauna.
Dr. Veyrenche is a clinical virologist working in the clinical microbiology department at Necker-Enfants Malades University Hospital in Paris, France. His research interests include the detection of emerging viral infection and clinical metagenomics.
Acknowledgment
Written informed consent for publication was obtained from the patient’s next of kin.
References
- Thibault PA, Watkinson RE, Moreira-Soto A, Drexler JF, Lee B. Zoonotic potential of emerging paramyxoviruses: knowns and unknowns. Adv Virus Res. 2017;98:1–55. DOIPubMedGoogle Scholar
- Diel DG, da Silva LHA, Liu H, Wang Z, Miller PJ, Afonso CL. Genetic diversity of avian paramyxovirus type 1: proposal for a unified nomenclature and classification system of Newcastle disease virus genotypes. Infect Genet Evol. 2012;12:1770–9. DOIPubMedGoogle Scholar
- Lu X, Wang X, Zhan T, Sun Y, Wang X, Xu N, et al. Surveillance of class I Newcastle disease virus at live bird markets and commercial poultry farms in eastern China reveals the epidemic characteristics. Virol Sin. 2021;36:818–22. DOIPubMedGoogle Scholar
- Dimitrov KM, Abolnik C, Afonso CL, Albina E, Bahl J, Berg M, et al. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infect Genet Evol. 2019;74:
103917 . DOIPubMedGoogle Scholar - Chong YL, Lam TTY, Kim O, Lu H, Dunn P, Poss M. Successful establishment and global dispersal of genotype VI avian paramyxovirus serotype 1 after cross species transmission. Infect Genet Evol. 2013;17:260–8. DOIPubMedGoogle Scholar
- Pestka D, Stenzel T, Koncicki A. Occurrence, characteristics and control of pigeon paramyxovirus type 1 in pigeons. Pol J Vet Sci. 2014;17:379–84. DOIPubMedGoogle Scholar
- Goebel SJ, Taylor J, Barr BC, Kiehn TE, Castro-Malaspina HR, Hedvat CV, et al. Isolation of avian paramyxovirus 1 from a patient with a lethal case of pneumonia. J Virol. 2007;81:12709–14. DOIPubMedGoogle Scholar
- Kuiken T, Breitbart M, Beer M, Grund C, Höper D, van den Hoogen B, et al. Zoonotic infection with pigeon paramyxovirus type 1 linked to fatal pneumonia. J Infect Dis. 2018;218:1037–44. DOIPubMedGoogle Scholar
- Zou X, Suo L, Wang Y, Cao H, Mu S, Wu C, et al. Concurrent pigeon paramyxovirus-1 and Acinetobacter baumannii infection in a fatal case of pneumonia. Emerg Microbes Infect. 2022;11:968–77. DOIPubMedGoogle Scholar
- Hurley S, Eden JS, Bingham J, Rodriguez M, Neave MJ, Johnson A, et al. Fatal human neurologic infection caused by pigeon avian paramyxovirus-1, Australia. Emerg Infect Dis. 2023;29:2482–7. DOIPubMedGoogle Scholar
- Winter S, Lechapt E, Gricourt G, N’debi M, Boddaert N, Moshous D, et al. Fatal encephalitis caused by Newcastle disease virus in a child. Acta Neuropathol. 2021;142:605–8. DOIPubMedGoogle Scholar
- Cui S, Xiong H, Feng Z, Chu Y, Que C, Qin J, et al. Severe pigeon paramyxovirus 1 infection in a human case with probable post-COVID-19 condition. Emerg Microbes Infect. 2023;12:
2251600 . DOIPubMedGoogle Scholar - World Organisation for Animal Health. Newcastle disease (infection with Newcastle disease virus). Chapter 3.3.10. In: Manual of diagnostic tests and vaccines for terrestrial animals. Paris : The Organisation; 2021 [cited 2026 Apr 27]. https://www.woah.org/fileadmin/Home/eng/Health_standards/tahm/3.03.10_NEWCASTLE_DIS.pdf
- Mallet F, Diouf L, Meunier B, Perret M, Reynier F, Leissner P, et al. Herpes DNAemia and TTV viraemia in intensive care unit critically ill patients: a single-centre prospective longitudinal study. Front Immunol. 2021;12:
698808 . DOIPubMedGoogle Scholar
Figures
Cite This ArticleOriginal Publication Date: April 28, 2026
1These authors contributed equally to this article.
2These senior authors contributed equally to this article.
Table of Contents – Volume 32, Number 5—May 2026
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Nolwenn M Dheilly, Institut Pasteur, Pathogen Discovery Laboratory, 25-28 rue du Dr. Roux, 75015, Paris, France
Top