Volume 32, Number 8—August 2026
Dispatch
Camel Prion Disease, Tataouine, Tunisia, 2019–2021
Cite This Article
Citation for Media
Abstract
We report 6 cases of camel prion disease in dromedaries in Tunisia, confirming widespread occurrence of the disease in North Africa. Affected animals showed neurologic signs and disease-associated prion protein accumulation in brain and lymphoid tissues. These findings highlight the importance of active surveillance and investigation of the epidemiology, transmission, and public health implications of this disease.
In 2018, a novel animal prion disease was identified in Algeria, termed camel prion disease (CPrD), affecting a previously unreported host species, the dromedary camel (Camelus dromedarius) (1). After identification of CPrD in Algeria, an epidemiologic surveillance network was set up in Tunisia to monitor neurologic diseases in dromedaries. We describe results of investigations of suspected cases and report the detection of 6 CPrD cases.
Tunisia hosts ≈57,000 dromedaries, 75% of them in the south. Accordingly, investigations focused on Tataouine, the southernmost governorate. Over 3 years, we identified 8 dromedary camels showing neurologic and behavioral signs consistent with CPrD (1), raising suspicion of prion disease (Table).
Figure 1
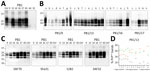
Figure 1. Detection and characterization of the proteinase K–resistant core (PrPres) of the pathological isoform of prion protein (PrPSc) from brain tissues of dromedary camels, Tunisia, 2019–2021. A)...
In addition to a histopathologic examination for spongiform changes, we examined all brains for the presence of the pathological isoform of prion protein (PrPSc) by using Western blot (WB) and immunohistochemistry (IHC) (Appendix). We performed WB analyses by using the TeSeE western blot kit (Bio-Rad Laboratories, https://www.bio-rad.com) and the ISS discriminatory WB method, validated for the animal prion diseases surveillance in Europe. Six animals tested positive, revealing the presence of the pathognomonic protease-resistant PrPSc (PrPres), characterized by the classical 3-band pattern in the 18–30 kDa range, in all available brain regions (Appendix Table 1), providing evidence of the involvement of several brain areas (Table; Figure 1, panels A, B). Conversely, 2 animals tested negative.
After confirmation of PrPSc presence, we conducted in-depth analysis of the 6 positive cases to investigate PrPres features, including protease cleavage site, presence of additional fragments, and glycosylation pattern. We treated samples with high concentrations of proteinase K to clearly define the cleavage site and probed them with a panel of monoclonal antibodies spanning the PrP sequence (Appendix Table 2, Figure 1).
PrPres from dromedary isolates displayed a higher apparent molecular weight of the unglycosylated band than the scrapie control, and we detected no additional C-terminal or internal fragments (Figure 1, panel C; Appendix Figure 1). PrPres exhibited lower overall glycosylation levels compared with scrapie, mainly in cortical areas relative to subcortical regions (Figure 1, panel D; Appendix Figure 2). The PrPres characteristics of the cases, including the electrophoretic profile, molecular weight and glycoprofile, are consistent with those previously reported in CPrD cases in Algeria (1).
We initially performed histopathologic and immunohistochemical analyses on the medulla oblongata and cerebellum, the primary target regions for animal TSE surveillance. We subsequently extended the analyses to other areas of the brains available (Appendix Table 1).
Figure 2
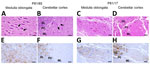
Figure 2. Histopathologic and immunohistochemical analyses of medulla oblongata and cerebellum of dromedary camels, Tunisia, 2019–2021. Images show the analyses performed on medulla oblongata (A, E, C, G) and cerebellum (B, F,...
Histopathologic examination revealed spongiform changes in only 2 animals (P81/16 and P81/65), in both obex (mainly in the dorsal vagal nucleus and the nucleus of the solitary tract) and cerebellar cortex (molecular layer), whereas we observed no lesions in the other animals (Table; Figure 2, panels A–D). Immunohistochemistry performed on the medulla oblongata and cerebellum demonstrated the presence of PrPSc in all samples except P81/15 and P81/64. In the obex, we observed intense PrPSc immunoreactivity in the dorsal vagal nucleus, the nucleus of the solitary tract (Figure 2, panels E, G), the hypoglossal and olivary nuclei, and the reticular formation. In the cerebellum, we detected extensive PrPSc immunolabeling throughout all layers of the cerebellar cortex (Figure 2, panels F, H) and in the white matter.
Figure 3
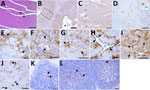
We extended histopathological and IHC analyses to other brain regions available for each case (Appendix Table 3). We observed mild spongiform changes exclusively in the cortices of animal P81/16 (Figure 3, panel A) and scattered vacuoles in the basal ganglia of P81/13 and P81/14.
IHC confirmed the diagnosis established from the medulla and cerebellum, identifying the same 6 CPrD-positive animals. In the cortex, PrPSc deposition was predominantly localized to layers I, V, and VI (Figure 3, panels B, C). The basal ganglia and thalamus (Figure 3, panel D) showed marked PrPSc accumulation in gray and white matter. We identified multiple PrPSc deposition patterns, including punctate and diffuse, intraneuronal, perineuronal, stellate, and perivascular (Figure 3, panels E–I). Of note, in the medulla oblongata, we observed a dense intra-astrocytic PrPSc deposition that filled the entire cytoplasm (Figure 3, panel J). IHC revealed PrPSc deposits in the primary and secondary follicles of all available lymph nodes, with variable immunostaining intensity (Table; Figure 3, panels K, L).
We conducted sequencing analysis of the entire PrP coding sequence. The analysis revealed that the 7 dromedary camels shared the same genetic background, being homozygous for the wild-type prion protein allele (Table).
In this study, we report the detection of CPrD in dromedary camels in Tunisia, providing further evidence for the geographic distribution of the disease and supporting the hypothesis of its broader presence in North Africa. Analyses were concordant across methods; 6 of 8 suspected camels tested positive, confirming that standard tools (e.g., kits, WB, IHC, monoclonal antibodies) reliably detect PrPSc in CPrD and provide effective methods for diagnosis and surveillance.
The detection of PrPSc in all available lymph nodes, together with the relatively young age of some camels, supports the hypothesis that CPrD is a transmissible prion disease. As in scrapie and chronic wasting disease, lymphoid involvement may enable extraneural propagation and serve as a source of environmental contamination (2–5), raising concerns about prion shedding and persistence.
Despite the typically long incubation periods of prion diseases, several affected camels were relatively young, showing clinical signs at 3 years of age, when sexual maturity is only just reached (3–5 years). That finding might reflect early-life or high-dose exposure, given that extensive environmental contamination can shorten incubation (6,7). Similar patterns are reported in scrapie and chronic wasting disease, where vertical and early-life transmission contribute to infection in young animals (4,8–14).
The recent detection of sheep scrapie in the same area of Tunisia as CPrD (15) raises the question of possible links, although no similarities have yet been shown. Bioassays will be crucial to clarify strain relationships and whether environmental factors might facilitate cross-species transmission or adaptation.
Taken together, the data from Algeria and Tunisia suggest that CPrD may be endemic in certain areas. Despite the limited number of reported cases so far, the absence of active surveillance programs in those countries, unlike in Europe for scrapie and bovine spongiform encephalopathy (BSE), raises the possibility of a higher number of undetected cases. Given frequent cross-border movements and the extensive pastoral systems in which camels roam widely, contact between herds in Tunisia and herds in Algeria is frequent and has possible implications for disease circulation.
CPrD may substantially affect camel health and production, given that dromedaries are long-lived animals with extended reproductive and productive roles and are vital to pastoralist communities in arid regions. Its detection in areas where camels are central to food production underscores the need to understand its epidemiology, transmission, and long-term impact.
The emergence of a new animal prion disease also raises public health concerns. Although only BSE has shown zoonotic potential, its history highlights the necessity of monitoring all animal prion diseases. Long incubation and absence of early clinical markers complicate risk assessment, as exemplified by the decade-long delay between cattle BSE and the variant Creutzfeldt–Jakob disease epidemic in humans (2).
Given the potential implications for animal and human health and the possible socioeconomic impact in regions where dromedaries play a key role, ongoing surveillance and in-depth characterization of CPrD are essential to enhance our comprehension of its epidemiology, host range, and zoonotic potential.
Prof. Amara is emeritus professor of histology and pathological anatomy at the National School of Veterinary Medicine of Sidi Thabet in Sidi Thabet, Tunisia. He is also vice president of the Expert Committee on Animal Health at the National Center for Veterinary Surveillance in Tunisia. His research focuses on prion diseases in ruminants and dromedaries.
Acknowledgments
We would like to thank Baaissa Babelhadj, who first described CPrD, for providing us with the samples from the dromedaries in Algeria. This contribution was essential because it enabled a direct comparison with the cases in Tunisia included in our study.
This study was supported by the Istituto Superiore di Sanità, Independent Research Program (grant no. ISS20-97e1d82bda5a) and by the La Sapienza University of Rome and Istituto Superiore di Sanità, Italy–Africa PhD Program (grant no. 5165/2023).
References
- Babelhadj B, Di Bari MA, Pirisinu L, Chiappini B, Gaouar SBS, Riccardi G, et al. Prion disease in dromedary camels, Algeria. Emerg Infect Dis. 2018;24:1029–36. DOIPubMedGoogle Scholar
- Houston F, Andréoletti O. Animal prion diseases: the risks to human health. Brain Pathol. 2019;29:248–62. DOIPubMedGoogle Scholar
- Moore SJ, Kunkle R, Greenlee MH, Nicholson E, Richt J, Hamir A, et al. Horizontal transmission of chronic wasting disease in reindeer. Emerg Infect Dis. 2016;22:2142–5. DOIPubMedGoogle Scholar
- Saunders SE, Bartelt-Hunt SL, Bartz JC. Occurrence, transmission, and zoonotic potential of chronic wasting disease. Emerg Infect Dis. 2012;18:369–76. DOIPubMedGoogle Scholar
- van Keulen LJ, Schreuder BE, Vromans ME, Langeveld JP, Smits MA. Pathogenesis of natural scrapie in sheep. Arch Virol Suppl. 2000;16:57–71.PubMedGoogle Scholar
- Fryer HR, McLean AR. There is no safe dose of prions. PLoS One. 2011;6:
e23664 . DOIPubMedGoogle Scholar - Ryder SJ, Dexter GE, Heasman L, Warner R, Moore SJ. Accumulation and dissemination of prion protein in experimental sheep scrapie in the natural host. BMC Vet Res. 2009;5:9. DOIPubMedGoogle Scholar
- Foster JD, Goldmann W, Hunter N. Evidence in sheep for pre-natal transmission of scrapie to lambs from infected mothers. PLoS One. 2013;8:
e79433 . DOIPubMedGoogle Scholar - Konold T, Moore SJ, Bellworthy SJ, Terry LA, Thorne L, Ramsay A, et al. Evidence of effective scrapie transmission via colostrum and milk in sheep. BMC Vet Res. 2013;9:99. DOIPubMedGoogle Scholar
- Lacroux C, Corbière F, Tabouret G, Lugan S, Costes P, Mathey J, et al. Dynamics and genetics of PrPSc placental accumulation in sheep. J Gen Virol. 2007;88:1056–61. DOIPubMedGoogle Scholar
- Nalls AV, McNulty E, Powers J, Seelig DM, Hoover C, Haley NJ, et al. Mother to offspring transmission of chronic wasting disease in reeves’ muntjac deer. PLoS One. 2013;8:
e71844 . DOIPubMedGoogle Scholar - Race R, Jenny A, Sutton D. Scrapie infectivity and proteinase K-resistant prion protein in sheep placenta, brain, spleen, and lymph node: implications for transmission and antemortem diagnosis. J Infect Dis. 1998;178:949–53. DOIPubMedGoogle Scholar
- Selariu A, Powers JG, Nalls A, Brandhuber M, Mayfield A, Fullaway S, et al. In utero transmission and tissue distribution of chronic wasting disease-associated prions in free-ranging Rocky Mountain elk. J Gen Virol. 2015;96:3444–55. DOIPubMedGoogle Scholar
- Spiropoulos J, Hawkins SA, Simmons MM, Bellworthy SJ. Evidence of in utero transmission of classical scrapie in sheep. J Virol. 2014;88:4591–4. DOIPubMedGoogle Scholar
- Amara A, Elmehatli K, Di Bari MA, Pirisinu L, Andolsi R, Gachout S, et al. Characterization of the first case of classical scrapie in a sheep in Tunisia. Transbound Emerg Dis. 2023;2023:
2253316 . DOIPubMedGoogle Scholar
Figures
Table
Cite This ArticleOriginal Publication Date: July 16, 2026
1These authors contributed equally to this article.
Table of Contents – Volume 32, Number 8—August 2026
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Laura Pirisinu, Department of Food Safety, Nutrition and Veterinary Public Health, Istituto Superiore di Sanità, Viale Regina Elena 299, 00161 Rome, Italy
Top