Volume 6, Number 4—August 2000
Perspective
Migratory Birds and Spread of West Nile Virus in the Western Hemisphere
Migratory Birds and West Nile Virus in the Old World
Migratory Birds and West Nile Virus in the New World
Normal Interhemispheric Migration
Displacement of West African Birds to the New World by Tropical Storms
Legal and Illegal Importations of Domestic Birds
Animals Other Than Birds and Entry of West Nile Virus into the New World
Migratory Birds and West Nile Virus Distribution in the Western Hemisphere
Bird Migration in the New York Region
Conclusions
Cite This Article
Cite This Article
Citation for Media
Abstract
West Nile virus, an Old World flavivirus related to St. Louis encephalitis virus, was first recorded in the New World during August 1999 in the borough of Queens, New York City. Through October 1999, 62 patients, 7 of whom died, had confirmed infections with the virus. Ornithophilic mosquitoes are the principal vectors of West Nile virus in the Old World, and birds of several species, chiefly migrants, appear to be the major introductory or amplifying hosts. If transovarial transmission or survival in overwintering mosquitoes were the principal means for its persistence, West Nile virus might not become established in the New World because of aggressive mosquito suppression campaigns conducted in the New York area. However, the pattern of outbreaks in southern Europe suggests that viremic migratory birds may also contribute to movement of the virus. If so, West Nile virus has the potential to cause outbreaks throughout both temperate and tropical regions of the Western Hemisphere.
The first known human case of West Nile virus infection recorded in the Western Hemisphere was reported in August 1999 (1). Eventually, 62 cases of the disease were confirmed; no new cases have been reported since October 16, 1999 (2). Of the human cases laboratory-confirmed as of October 8, 1999, 70% occurred within a circle 10 km in radius centered in the northern end of the New York City borough of Queens (1). Coincident in both space and time with the human outbreak was a substantial die-off of birds, particularly American Crows (Corvus brachyrhynchos) (3). Unusual bird deaths were noted around the Bronx Zoo in mid-August (4), approximately 8 km north of the epicenter of the human epidemic. Several thousand crows and other avian species are presumed to have died of the virus, mostly in and around the New York City area (5). In addition to wild birds, the die-off included specimens in the Bronx and Queens zoo collections (including Chilean Flamingos [Phoenicopterus chilensis], Guanay Cormorants [Phalacrocorax bougainvillei], Bald Eagles [Haliaeetus leucocephalus], Black-billed Magpies [Pica pica], Bronze-winged Ducks [Anas specularis], Impeyan Pheasants [Lophophorus impeyanus], Blyth's Tragopan [Tragopan blythi], and Snowy Owls [Nyctea scandia]) (6).
The spatial and temporal juxtaposition of avian and human infections in this instance and historically has led many epidemiologists to conclude that birds act as introductory hosts, perhaps by infecting ornithophilic mosquitoes, which in turn infect amplifying hosts and eventually humans (3,7). Despite the fact that migratory birds have long been suspected as critical agents in outbreaks of this and other arboviruses, the link remains conjectural because of the difficulty in determining the intensity and duration of viremia in naturally infected wild birds (8,9).
We present an overview of the association of West Nile virus with birds, focusing in particular on the advent and movement of the virus in the Western Hemisphere.
West Nile virus was first isolated and identified as a distinct pathogen from the blood of a woman in the West Nile region of Uganda in 1937 (10). Cross-neutralization tests have been used to classify the virus as a Flavivirus (Family Flaviviridae) closely related to Japanese encephalitis virus, eastern Asia; Kunjin virus, Australia and Southeast Asia; and St. Louis encephalitis virus, North and South America (11,12). Several genetic lineages of the virus have been identified in different geographic locations. The lineage associated with the New York outbreak has been identified as virtually identical with an Israeli strain of the virus by phylogenetic analysis of E-glycoprotein nucleic acid sequence data (13). West Nile virus was one of the most widespread flaviviruses, even before its recent entry into North America. Its distribution extends throughout Africa, the Middle East, and southern temperate and tropical Eurasia (14,15). During the 1950s, an estimated 40% of the human population in Egypt's Nile Delta was seropositive for the virus (16). The largest human epidemic occurred in Cape Province, South Africa, in 1974, when approximately 3,000 clinical cases of the virus were recorded (17).
Migratory birds have long been suspected as the principal introductory hosts of West Nile virus into new regions for the following reasons: Outbreaks of the virus in temperate regions generally occur during late summer or early fall, coinciding with the arrival of large concentrations of migratory birds (and mosquitoes) (18-20); these outbreaks often occur among humans living in or near wetlands where high concentrations of birds come into contact with large numbers of ornithophilic mosquitoes (7,21,22); the principal vectors from which the virus has been isolated are mainly ornithophilic mosquitoes (Culex univittatus in the Middle East and C. pipiens in Europe) (7,13,14,18,22); antibodies to the virus have been found in the blood of many migratory bird species in Eurasia (14,15,22-24); migratory birds have been linked with transporting related viruses in the Western Hemisphere (9,25,26); West Nile virus has been isolated from some species of actively migrating birds (e.g., the Barred Warbler [Sylvia risoria] in Cyprus and the Turtle Dove [Streptopelia turtur] in Slovakia (20,23,24); viremia sufficiently long-term to infect vector mosquitoes has been documented in several bird species (18,19,27-29); and migration places substantial physiologic stress on birds. Stress has been shown to promote immunosuppression and enhanced replication of West Nile virus in rodents (30).
Further support for the possibility that migratory birds play a major role in virus transport comes from study of related viruses. For instance, both Eastern (EEE) and Western Equine Encephalomyelitis alphaviruses, ecologic relatives of West Nile virus, have been isolated from actively migrating birds in the United States (25,26). Evidence also indicates that the 1962 epidemic of EEE in Jamaica resulted from transport of the virus by birds from the continental United States (31).
Unlike the 1999 New York City epidemic, during which large numbers of dead and dying birds, especially crows, were observed concurrently with clinical reports of human infection with the virus (3), the Old World epidemics of West Nile virus had few concurrent reports of deaths of infected birds (7). This difference could indicate lack of both exposure and adaptation to the virus among New World avian populations compared with Old World species. Old World data indicate that susceptibility to fatal infection with the virus varies markedly for adult and young birds, with high death rates in juvenile and high incidence of circulating antibodies in adult birds (19). Susceptibility to infection also varies considerably among species. Hooded Crows (Corvus corone) had both a high death rate in young birds in laboratory experiments and high levels of circulating antibodies in adults, while Rock Doves (Columba livia) appeared to be much less susceptible to both infection and death from the virus (19).
As was the case for humans, the first birds documented as infected with West Nile virus in the Western Hemisphere were identified in August 1999 (6). Thereafter, large die-offs of wild and captive birds at the Bronx Zoo and other parts of the New York area coincided with the increasing number of human cases reported from the same region (1,4,6). As in several European outbreaks, the main vector in the New York City epidemic was identified as the ornithophilic mosquito C. pipiens (4). Furthermore, the outbreak in humans occurred at urban sites near wetlands where migratory birds, ornithophilic mosquitoes, and humans were concentrated. These circumstances, in conjunction with the ecology of the virus in the Old World, support the conjecture that zoo, pet, domestic, or wild birds were responsible for introducing the virus to the New World. If so, birds could have served as the source by normal migration, displacement from normal range by storms, or importation (legal and illegal).
A small percentage of the populations of a few bird species migrate regularly in August and September from breeding grounds in the Old World to wintering grounds along the eastern seaboard of North America. An example of this group is the Eurasian Wigeon (Anas penelope), which breeds across the entire Palearctic region from Iceland to Siberia's Kamchatka Peninsula and winters primarily in the temperate and tropical zones of the Old World (32), where contact with West Nile virus is possible. However, a few Eurasian Wigeons, presumably from the Icelandic breeding population, winter regularly along the coast of eastern North America (32,33). These birds could contract the virus from vector mosquitoes infected by biting other members of the breeding population that winter in areas where West Nile virus is prevalent and bring infectious blood to the New World on their winter migrations.
Eurasian Wigeons are not the only species with such a migration pattern. Eurasian populations of several species in which evidence of exposure (e.g., antibodies) to the virus has been detected are rare migrants along the eastern seaboard of North America (Table 1). However, if normal migration were a likely pathway, the virus would likely have become established earlier in this hemisphere, since individual birds of several species known to be susceptible to the virus migrate annually from Eurasia to the United States (Table 1). However, the numbers of migrants are so small that the probability of the cooccurrence of an infectious migrant, ornithophilic vector mosquitoes, and numerous avian amplifying hosts seems low. Furthermore, the most likely form of the virus carried by migrants would be that from West Africa, because that is where most western European-breeding populations of these species winter. The New York City strain of the virus was nearly identical to that found in the Middle East, which is different from the West African strain (13). Despite these considerations, normal migration remains a distinct possibility as the mode of entry for the disease.
A very few birds, particularly seabirds, are carried by tropical storms across the Atlantic each summer from their normal environs on or near the coast of West Africa (39). A number of such storms form each summer and fall near the Cape Verde Islands off the western coast of Africa, travel across the Atlantic, and occasionally reach land the east coast of North America, depositing birds that were carried thousands of kilometers from their homes. Species known to have been infected by West Nile virus and whose habitat and distribution indicate that they might be affected by such displacement include the Gray Heron (Ardea cinerea), the Little Egret (Egretta garzetta), the Cattle Egret (Bubulcus ibis), the Black-headed Gull (Larus ridibundus), and the Yellow-legged Gull (Larus cachinnans) (Table 1). The same objections apply to this scenario for the introduction of the virus to the New World as for normal migration, i.e., low numbers and the likelihood that a storm-transported bird would be infected with the West African rather than the Middle Eastern form of the virus.
Although the legal importation of pet, zoo, and domestic birds (e.g., geese, ducks, turkeys, and chickens) has declined since enactment of the 1992 Wild Bird Conservation Act, 2,770 birds entered the country through John F. Kennedy International Airport in 1999: 323 pet birds and 2,447 commercial birds; an additional 12,931 birds passed through in transit (S. Kaman, USDA, Pers. Comm.). All legal importations are subject to U.S. Department of Agriculture (USDA/Animal and Plant Health Inspection Service) and U.S. Fish and Wildlife Service rules, regulations and procedures, and most birds undergo a quarantine of at least 30 days at USDA facilities located in the vicinity of three U.S. ports-of-entry (New York, Los Angeles, Miami). During quarantine the birds are isolated indoors in air-filtered isolation cages to prevent the transmission of communicable diseases, medicated for psittacosis, and tested for diseases affecting poultry (e.g., Newcastle Disease) (40). Even with such precautions, some birds infected with West Nile virus could be bitten by mosquitoes during transit to quarantine or could escape detection during quarantine; no tests specific for the virus are performed, and many Old World species remain asymptomatic if infected. After release from quarantine, viremic birds could transmit the virus to native birds if held outdoors in habitats (e.g., zoos) supporting both ornithophilic mosquitoes and concentrations of birds. Such a scenario is even more probable in the case of illegally imported birds, which would not be subject to quarantine or even cursory health examinations.
Although birds appear to be the principal means by which the virus moves from site to site in the Old World, other modes of entry into the New World are possible. Humans, horses, and some other mammals are highly susceptible to infection by the virus (7), and not all become too sick to travel during periods of potential viremia. Furthermore, it is unlikely that all animal hosts and vectors have been identified. Intercontinental air travel could transport infectious animals from viral foci in Eurasia or Africa. An infectious mosquito may enter a plane in one of these areas, travel to New York, and infect a person, horse, or bird en route or after arrival.
Since identification of West Nile virus in the United States, a massive effort has been undertaken to determine when and where the virus will next appear in the human population. The New York City area is thought most likely to be affected by the next epidemic. If transovarial transmission in mosquitoes or survival in overwintering mosquitoes were the principal means for persistence of this virus, annual recurrence of the virus might be expected at the same sites. However, while transovarial transmission of the virus has been documented, it occurs at low levels (< 1%) (15,41). In addition, public health officials have implemented mosquito-control measures that are likely to continue. Furthermore, although the virus may remain enzootic for years awaiting environmental conditions that favor an epidemic (36), the usual pattern in southern Eurasia is one of isolated outbreaks, apparently resulting from importation of active virus by migratory birds into an area with appropriate climatic, vector, and amplifying host conditions (7). This pattern suggests that future movements of the virus in the Western Hemisphere may depend on its persistent amplification in wintering avian populations in New World tropical, subtropical, and southern temperate regions, and subsequent importation into major avian concentration areas in the temperate region.
If this Old World pattern persists, the New York area is unlikely to be the site of the next human outbreak because the occurrence of optimal combinations of infecting host, vector, amplifying host, and susceptible human population depends on substantial annual variation based on stochastic environmental factors (e.g., rainfall and temperature) (7). The known ecology of West Nile virus indicates that the virus is more likely to persist in the Western Hemisphere if it is translocated by avian hosts to southern wintering sites. Old World data indicate that ideal over-wintering conditions for West Nile virus combine three key factors: a viremic, infectious host bird; active, ornithophilic mosquitoes to serve as vectors; and large numbers of one or more amplifying avian host species. This combination of numerous wintering birds and ornithophilic mosquitoes (e.g., in southern wetlands or wet agricultural or urban areas) could provide amplification and a permanent base for the virus from which it could be spread northward by migrating birds. Thus, understanding the major migration patterns of the potential infecting host species through the New York City region may hold the key to understanding the future of the virus in the Hemisphere.
Four major routes are followed by birds that gather in and pass through the New York City area (defined as a circle 10 km in radius centered on north Queens) in late summer and early fall: the southeastern U.S. route, the circum-Gulf route, the trans-Gulf route, and the Caribbean island-western North Atlantic route.
Southeastern U.S. Route
Figure 1

Members of approximately 155 species of birds may originate in or pass through the New York area on their way to wintering grounds in the southeastern United States, following the route described by W.W. Cooke for the American Robin (Turdus migratorius) (42). Most of these species are not suitable hosts from the perspective of the virus, because they follow individual migration paths and do not gather in large numbers in habitats with high concentrations of ornithophilic mosquitoes. However, approximately 32 species of birds that follow the southeastern U.S. migration route would be likely to occur in high densities in or near wetlands on migration as they pass through New York and on to their wintering sites (Table 2). The European Starling (Sturnus vulgaris), normally considered to be "sedentary," is an example of a species a portion of whose northeastern populations follows such a migratory route (Figure 1). American Crows are another such species, as well as the Mallard (Anas platyrhynchos) (39). Mallards breed throughout the temperate and boreal regions of North America and, in late summer, members of the eastern Canadian populations begin to collect in flocks at wetlands preparatory for migration to wintering sites in the southeastern United States. Many Mallards from the northeastern United States also migrate to the southeastern United States, although some of the birds remain at their breeding grounds throughout the winter. Mallards migrating south gather in flocks along the migration route, as well as on the wintering grounds. However, between stop-over sites, each bird follows its own migration path, so flock membership is changing continuously. As a result, birds found together as transients at a pond in Long Island or the Bronx are not likely to be members of the same winter flock in coastal Georgia or Alabama. After the wintering site has been reached or during northward movement in the spring, males search for mates. Once paired, a male follows his mate back to her natal area, which ensures wide dispersal of male birds throughout the Mallard's entire North American breeding range, a pattern seen in many duck species (43).
Circum-Gulf Route
Figure 2

Birds of some species that typically winter in Mexico and Central America avoid crossing large expanses of open water; these species include hawks, herons, and egrets, as well as some ducks and gulls. Although some birds may fly directly across the Gulf of Mexico to their winter quarters, others detour around the Gulf, passing along its western coast during both south- and northward migratory journeys. Approximately 11 of these species might flock together in and around New York wetlands during migration and again on wintering grounds (Table 2). An example is the Herring Gull (Larus argentatus) (Figure 2).
Trans-Gulf Route
Approximately 125 species of birds have populations that transit the New York region on their way to wintering grounds in Mexico and Central America following a trans-Gulf route in the fall (42,44). Distributional data indicate that most do not follow the same route north in the spring, but instead take a more westerly route over or paralleling the western Gulf coast (45). Also, many birds of these species, especially young birds or those from populations in the center of the continent, follow a circum-Gulf route both south and north. Although this group includes many migratory birds that breed in eastern North America, only approximately 12 species (Table 2) could serve as carriers for West Nile virus, because most migrate alone and do not gather in large roosting or feeding flocks in the winter or during migration (46).
Caribbean Island/Western North Atlantic Route
Figure 3
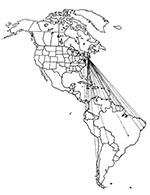
Approximately 70 species of birds have populations that pass through New York and cross the western North Atlantic or Caribbean Sea en route south to their wintering grounds on Caribbean islands or in South America. Like the trans-Gulf pattern, this route is elliptical, with birds following a more westerly route across the Gulf of Mexico or along its western shore in the spring (44-46). Members of approximately 22 species of birds that gather in flocks during migration and during the winter follow this route (Table 2) (e.g., the Common Tern [Sterna hirundo]) (Figure 3).
Members of one or more avian species that pass through New York and gather in wetlands in large, dense groups potentially reach every part of the southeastern United States, Mexico and Central America, the Caribbean Islands, and South America during their migration south to wintering sites and nearly every part of North America during their migration north to breeding sites (Table 2, Figures 1-3).
Movement of the virus in the Old World seems to involve a set of conditions including infectious avian host, numerous ornithophilic mosquito vectors, and cross-species transmission to a numerous avian amplifying host (not necessarily the same species as the infectious host but the same location). Therefore, because outbreaks are dependent on a series of probabilities, the suitable wetland sites likely to receive the largest number of potentially infected hosts seem to be the most likely place for future outbreaks. Banding data, showing winter concentration areas for large numbers of birds (Table 2), indicate that the most likely place for a bird-spawned outbreak on the wintering ground would be along the coastal plain of Georgia, northern Florida, or Alabama—if the necessary ornithophilic mosquitoes are sufficiently active and abundant.
This information suggests that the best approach to minimizing effects of the virus on humans should involve intensive monitoring of fall and winter avian concentrations for abnormal die-offs; collecting and testing bird carcasses from such die-offs; and implementation of mosquito control measures at die-off sites. In addition, aviculturalists, poultry markets, zoos, and others involved with shipment of birds from one part of the country to another should test birds for West Nile virus during quarantine to ensure that they are not bringing in or sending out infectious birds (47).
Future research should define criteria for predicting where the virus will go next. Defining the duration of viremia or the frequency of cycling of active virus in the various mosquito and avian populations that have been exposed to the virus is critical. At present, nothing is known about long-term persistence of active virus in the blood of New World avian species, and little is known about long-term viremia in other avian species. As noted by Blaskovic and Ernek (48), "The role of birds in ecology of arboviruses depends upon whether the migrating vector finds favourable conditions in the new environment and whether the local vectors are capable of transmitting the appropriate virus. The presence of antibody for arboviruses in migratory birds indicates only a virus-host interaction but does not explain when and where the infection occurred." In addition, detection of antibodies for West Nile virus in avian serum does not show whether the bird could serve as a source for transmission of active virus by mosquitoes to humans. Determining the viremic potential of North American bird species is critical to understanding the future of the virus in the New World and should be the focus of future research.
John Rappole is a research scientist at the Smithsonian Conservation and Research Center in Front Royal, Virginia, USA. He is interested in the ecology of migratory birds.
Acknowledgment
We thank W. C. Reeves for extensive comments and suggestions for improving the manuscript.
References
- Centers for Disease Control and Prevention. Update: West Nile-like viral encephalitis—New York, 1999. MMWR Morb Mortal Wkly Rep. 1999;48:890–2.PubMedGoogle Scholar
- Centers for Disease Control and Prevention. Guidelines for surveillance, prevention, and control of West Nile virus infection—United States, 2000. MMWR Morb Mortal Wkly Rep. 2000;49:25–8.PubMedGoogle Scholar
- Anderson JF, Andreadis TG, Vossbrinck CR, Tirrell S, Wakem EM, French RA, Isolation of West Nile virus from mosquitoes, crows, and a Cooper's Hawk in Connecticut. Science. 1999;286:2331–3. DOIPubMedGoogle Scholar
- Centers for Disease Control and Prevention. Outbreak of West Nile-like viral encephalitis—New York, 1999. MMWR Morb Mortal Wkly Rep. 1999;48:845–9.PubMedGoogle Scholar
- Rocke T. President's corner. [Suppl.]. J Wildl Dis. 2000;:36.
- Steele KE, Linn MJ, Schoepp RJ, Komar N, Geisbert TW, Manduca RM, Pathology of fatal West Nile virus infections in native and exotic birds during the 1999 outbreak in New York City. Vet Pathol. 2000;37:208–24. DOIPubMedGoogle Scholar
- Hubálek Z, Halouzka J. West Nile fever—a reemerging mosquito-borne viral disease in Europe. Emerg Infect Dis. 1999;5:643–50. DOIPubMedGoogle Scholar
- Reeves WC, Hammon WM. Epidemiology of the arthropod-borne viral encephalitides in Kern County, California, 1943-1952. University of California Publications in Public Health. Vol 4. Berkeley (CA): University of California Press; 1962.
- Smithburn KC, Hughes TP, Burke AW, Paul JH. A neurotropic virus isolated from the blood of a native of Uganda. Am J Trop Med Hyg. 1940;20:471–92.
- Calisher CH, Karabatsos N, Dalrymple JM, Shope RE, Porterfield JS, Westaway EG, Antigenic relationships between flaviviruses as determined by cross-neutralization tests with polyclonal antisera. J Gen Virol. 1989;70:37–43. DOIPubMedGoogle Scholar
- Wengler G, Bradley DW, Collett MS, Heinz FX, Schlesinger RW, Strauss JH. Family flaviviridae. VIth International Committee on Taxonomy of Viruses Report; 1999.
- Lanciotti RS, Roehrig JT, Deubel V, Smith J, Parker M, Steele K, Origin of the West Nile virus responsible for an outbreak of encephalitis in the northeastern United States. Science. 1999;286:2333–7. DOIPubMedGoogle Scholar
- Karabatsos N, ed. International catalogue of arboviruses, including certain other viruses of vertebrates. 3rd ed., and Supplements 1986-98. San Antonio (TX): American Society of Tropical Medicine and Hygiene; 1985.
- Peiris JSM, Amerasinghe FP. West Nile fever. In: Beran GW, Steele JH, editors. Handbook of zoonoses. 2nd ed. Section B: Viral. Boca Raton (FL): CRC Press; 1994. p. 139-48.
- Smithburn KC, Taylor RM, Rizk F, Kader A. Immunity to certain arthropod-borne viruses among indigenous residents of Egypt. Am J Trop Med Hyg. 1954;3:9–18.PubMedGoogle Scholar
- McIntosh BM, Jupp PG, Dos Santos I, Meenehan GM. Epidemics of West Nile and Sindbis viruses in South Africa with Culex (Culex) univittatus Theobald as vector. S Afr J Sci. 1976;72:295–300.
- Taylor RM, Work TH, Hurlbut HS, Rizk F. A study of the ecology of West Nile virus in Egypt. Am J Trop Med Hyg. 1956;5:579–620.PubMedGoogle Scholar
- Work TH, Hurlbut HS, Taylor RM. Indigenous wild birds of the Nile Delta as potential West Nile virus circulating reservoirs. Am J Trop Med Hyg. 1955;4:872–8.PubMedGoogle Scholar
- Nir Y, Goldwasser R, Lasowski Y, Avivi A. Isolation of arboviruses from wild birds in Israel. Am J Epidemiol. 1967;86:372–8.PubMedGoogle Scholar
- Lvov DK, Klimenko SM, Gaidamovich SY. Arboviruses and arbovirus infections (in Russian). Medicina (Moskva) 1989:1-335.
- Hubálek Z, Halouzka J. Arthropod-borne viruses of vertebrates in Europe. Acta Scientiarum Naturalium Brno. 1996;30:1–95.
- Watson GE, Shope RE, Kaiser MN. An ectoparasite and virus survey of migratory birds in the eastern Mediterranean. In: Cherepanov IA, editor. Transcontinental connections of migratory birds and their role in the distribution of arboviruses. Novosibirsk, Russia: Nauka; 1972. p. 176-80.
- Ernek E, Kozuch O, Nosek J, Teplan J, Folk C. Arboviruses in birds captured in Slovakia. Journal of Hygiene. Epidemiology and Immunology (Prague). 1977;21:353–9.
- Calisher CH, Maness KSC, Lord RD, Coleman PH. Identification of two South American strains of eastern equine encephalomyelitis virus from migrant birds captured on the Mississippi Delta. Am J Epidemiol. 1971;94:172–8.PubMedGoogle Scholar
- Lord RD, Calisher CH. Further evidence of southward transport of arboviruses by migratory birds. Am J Epidemiol. 1970;92:73–8.PubMedGoogle Scholar
- Semenov BF, Chunikhin SP, Karmysheva VY, Yakovleva NI. Studies of chronic arbovirus infections in birds. 1. Experiments with West Nile, Sindbis, Bhanja and SFS viruses [in Russian]. Vestnik Akademii Medicinskikh NaukSSSR (Moskva). 1973;2:79–83.
- Fedorova TN, Stavskiy AV. Latent infection of wild ducks with Omsk hemorrhagic fever and West Nile viruses (in Russian). In: Chumakov MP, editor. Aktualnye prolemy virusologii i profilaktiki virusnykh zabolevaniy. Moskva: Inst Poliomiel Virus Enc;1972. p. 226.
- Chunikhin SP. Introduction to arbovirus ecology [in Russian]. Med Virusol (Moskva). 1973;21:7–88.
- Ben-Nathan D. Stress and infectious disease. Israeli Journal of Veterinary Medicine. 1994;49:105–12.
- Work TH, Lord RD. Trans-gulf migrants and the epizootiology of arboviruses in North America. In: Cherepanov IA, editor. Transcontinental connections of migratory birds and their role in the distribution of arboviruses. Novosibirsk, Russia: Nauka; 1972. p. 207-14.
- Palmer RS, ed. Handbook of North American birds. Vol 2. New Haven (CT): Yale University Press; 1976.
- American Ornithologists' Union. Check-list of North American birds. 7th ed. Washington: The Union; 1998.
- Antipa C, Girjabu E, Iftimovici R, Draganescu N. Serological investigations concerning the presence of antibodies to arboviruses in wild birds. Revue Roumaine de Medecine. Serie de Virologie (Bucuresti). 1984;35:5–9.
- Taufflieb R, Robin Y, Bres P. Preliminary findings on the role of birds in the ecology of arboviruses in Senegal. In: Cherepanov IA, editor. Transcontinental connections of migratory birds and their role in the distribution of arboviruses. Novosibirsk: Nauka; 1972. p. 296-302.
- Hannoun C, Corniou B, Mouchet J. Role of migrating birds in arbovirus transfer between Africa and Europe. In: Cherepanov IA, editor. Transcontinental connections of migratory birds and their role in the distribution of arboviruses. Novosibirsk, Russia: Nauka; 1972. p. 167-72.
- Bull J. Birds of New York State. Garden City (NY): Doubleday; 1974.
- Work TH, Hurlbut HS, Taylor RM. Isolation of West Nile virus from hooded crow and rock pigeon in the Nile Delta. Proc Soc Exp Biol Med. 1953;84:719–22.PubMedGoogle Scholar
- Hurricane/tropical data for Atlantic. Unisys Weather. 1999. Available from: URL: http://weather.unisys.com/hurricane/atlantic/1999/index.html
- Importing a pet bird. United States Department of Agriculture, Animal and Plant Health Inspection Service. 2000. Available from: URL: http://www.aphis.usda.gov/oa/petbird.html
- Baqar S, Hayes CG, Murphy JR, Watts DM. Vertical transmission of West Nile virus by Culex and Aedes species mosquitoes. Am J Trop Med Hyg. 1993;48:757–62.PubMedGoogle Scholar
- Cooke WW. Bird migration. Washington. United States Department of Agriculture Bulletin. 1915;185:1–48.
- Bellrose FC. Ducks, geese and swans of North America. Harrisburg (PA): Stackpole Books; 1976.
- Rappole JH, Ramos MA, Oehlenschlager RJ, Warner DW, Barkan CP. Timing of migration and route selection in North American songbirds. In: Drawe L, editor. Proceedings of the First Welder Wildlife Foundation Symposium. Sinton (TX): Welder Wildlife Foundation; 1979. p. 199-214.
- Rappole JH, Ramos MA. Factors affecting migratory bird routes over the Gulf of Mexico. Bird Conserv Int. 1994;4:131–42. DOIGoogle Scholar
- Rappole JH. The ecology of migrant birds: a neotropical perspective. Washington: Smithsonian Institution Press; 1995.
- Snyder NFR, Derrickson SR, Beissinger SR, Wiley JW, Smith TB, Toone WD, Limitations of captive breeding in endangered species recovery. Conserv Biol. 1996;10:338–48. DOIGoogle Scholar
- Blaskovic D, Ernek E. Birds as hosts of arboviruses in Europe. In: Cherepanov IA, editor. Transcontinental connections of migratory birds and their role in the distribution of arboviruses. Novosibirsk, Russia: Nauka; 1972. p. 161-7.
Figures
Tables
Cite This ArticleTable of Contents – Volume 6, Number 4—August 2000
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
John H. Rappole, 1500 Remount Road, Front Royal, VA 22630, USA; fax: 540-635-6551
Top