Volume 14, Number 11—November 2008
Dispatch
Use of Malaria Rapid Diagnostic Test to Identify Plasmodium knowlesi Infection
Cite This Article
Citation for Media
Abstract
Reports of human infection with Plasmodium knowlesi, a monkey malaria, suggest that it and other nonhuman malaria species may be an emerging health problem. We report the use of a rapid test to supplement microscopic analysis in distinguishing the 5 malaria species that infect humans.
Recent reports of Plasmodium knowlesi infections in humans in Sarawak and Sabah in Borneo and in the Pahang Peninsula of Malaysia have focused attention on the potential of monkey malarias to be a human health issue (1,2). As much as 70% of malaria infections in regional hospitals in Borneo are the result of P. knowlesi infection; similar infections have been found in Thailand, the Philippines, and Singapore (3–5). To date, only patients in hospitals are being screened for the disease. To better understand the epidemiology of this apparent outbreak of P. knowlesi in humans, one needs a method to rapidly screen both monkeys and humans in areas of high disease prevalence, regardless of their present health status. Thus, a rapid test that could detect and distinguish among the primate malarias would not only benefit individual patients but would also provide an important epidemiologic tool to monitor the overall risk and prevalence of malaria.
We have known for nearly 8 decades that, under laboratory conditions, several monkey malarias are capable of infecting humans and that P. knowlesi can be transmitted to humans by mosquito bite (6,7). Work in Malaysia by a team from the National Institutes of Health nearly 50 years ago reported that transmission to humans was not occurring to any prevalent extent. Currently, we see major foci of the disease, which can be life-threatening. Although the current overall incidence of P. knowlesi infection in humans is low, an exacerbating problem is that it can be consistently misdiagnosed by microscopy as the more benign human malaria, P. malariae (1,2,8,9). The rapid replication rate of P. knowlesi and the resulting high level of parasitemia warrant immediate and aggressive treatment, whereas P. malariae does not. Although the use of PCR has been essential to defining the problem, a more rapid diagnosis would be an important tool for prompt medical treatment. Furthermore, incorporating the capability to detect P. knowlesi into existing rapid tests already capable of detecting the other 4 Plasmodium species that infect humans (P. falciparum, P. vivax, P. ovale, and P. malariae) would be beneficial.
P. knowlesi is transmitted by members of the Anopheles leucosphyrus group of mosquitoes that resides in the upper canopy of the forests in large areas of Southeast Asia; these Anopheles mosquitoes have infrequent contact with humans (10). With increasing encroachment into the forest areas to provide farmland, however, humans are likely to increase their exposure to this vector. The potential for P. knowlesi infection as well as other monkey malarias to expand into the human population is real. While the P. knowlesi parasite is carried by zoophilic mosquitoes, some monkey malarias such as P. cynomolgi and P. inui are transmitted by the same mosquito vectors that carry human malaria and therefore represent an even wider threat.
Figure 1
Figure 1. Binding specificity of different anti–Plasmodium lactate dehydrogenase (pLDH) antibodies. A) Shown are the reactivities of the indicated monoclonal antibodies (MAbs) to the LDH from 7 Plasmodium spp. Reactivity was determined by...
One important test developed for detecting human malarias is an antigen-capture test based on monoclonal antibodies (MAbs) to plasmodium lactate dehydrogenase (pLDH). The 4 human malarial LDH isoforms have been cloned, and >20 MAb have been raised that differentially recognize epitopes among the isoforms (11,12). The specificity of a subset of these antibodies is shown in Figure 1. Of the 4 human Plasmodium spp., antibodies such as 17E4 and 7G9 specifically bind only to P. falciparum LDH, whereas antibodies such as 11D9 and 13H11 bind only to P. vivax LDH.
Using this panel of antibodies, we show that we can distinguish P. knowlesi from P. malariae. P. knowlesi binds to both the “falciparum-specific” (17E4/7G9) and the “vivax-specific” (11D9/13H11) antibodies (Figure 1, panels A and B). Furthermore, P. knowlesi does not react with 10D12 (an antibody specific for P. ovale), 7E7 (an antibody that reacts strongly with P. malariae and weakly with P. falciparum), or 9C1 (an antibody that reacts exclusively with P. ovale and P. malariae). Detecting P. knowlesi in monkeys, which often are co-infected with several other malaria parasites, is also important and can be achieved with the same panel of antibodies. We have tested the reactivity of the P. falciparum–specific antibody (17E4/7G9) with the other monkey malarias known to be indigenous to Malaysia (P. cynomolgi, P. inui, and P. fieldi) and found that none react (Figure 1, panel C). This then serves as a basis for distinguishing P. knowlesi from the other prevalent forms of monkey malaria.
Figure 2
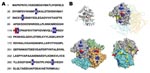
Figure 2. Modeling of the analysis of Plasmodium knowlesi lactate dehydrogenase (LDH). A) Sequence of LDH from P. knowlesi deduced from genomic DNA fragments sequenced by the Sanger malaria genome project (www.sanger.ac.uk/Projects/P_knowlesi)....
The unexpected pattern of antibody recognition on which we based our tests led us to examine the molecular basis of recognition (Figure 2). As expected, P. knowlesi LDH is highly similar to the known pLDH isoforms. We found that only a few residue differences could account for the epitope differences detected by the 17E4/7G9 and 11D9/13H11 antibodies. We first generated a 3-dimensional model of P. knowlesi LDH and then mapped surface-exposed residues that were uniquely shared by P. falciparum or P. vivax isoforms. The protein structure was calculated by using the structures of P. falciparum and P. vivax LDH (PDB: 2A94 and 2AA3) and the WURST threading server (13). Here, only a few patches of residues were found to describe the P. vivax–specific epitope, and only 1 residue (K115) was found to describe the P. falciparum–specific epitope (Figure 2, panel B). Thus, the existing MAbs perform well at distinguishing pLDH isoforms despite only a small number of different surface-exposed residues.
These data show that pLDH antibodies that detect P. falciparum and P. vivax can also be used to detect and distinguish P. knowlesi. The 1 major caveat is that a P. knowlesi infection cannot be distinguished from a mixed infection with both P. vivax and P. falciparum in the blood. Mixed infections of this description, however, are infrequent, as these species do not proliferate concurrently when both are present in the blood (14,15). Furthermore, any confusion would be resolved by microscopic examination of blood that, while inadequate to distinguish P. knowlesi and P. malariae, would serve to distinguish P. knowlesi from mixed infections.
Obviously, an antibody specific for P. knowlesi would be optimal if the threat of P. knowlesi increases. Although development of a specific antibody would be a considerable investment, our epitope analysis discussed here indicates that only small sequence differences in pLDH isoforms are required to generate antibody panels capable of uniquely distinguishing animal pLDH isoforms.
Dr McCutchan is head of the Regulation of Growth and Development Section of the Laboratory of Malaria and Vector Research at NIAID. His work on malaria, which spans over 25 years, has resulted in the publication of >100 manuscripts on such subjects as vaccine development, parasite diagnosis, and antimalaria drug development.
Acknowledgments
We thank Brenda Rae Marshall for editorial assistance.
This work was supported by the Intramural Research Program of the Division of Intramural Research, National Institute of Allergy and Infectious Diseases (NIAID), National Institutes of Health.
References
- Singh B, Kim Sung L, Matusop A, Radhakrishnan A, Shamsul SS, Cox-Singh J, A large focus of naturally acquired Plasmodium knowlesi infections in human beings. Lancet. 2004;363:1017–24. DOIPubMedGoogle Scholar
- Cox-Singh J, Davis TM, Lee KS, Shamsul SS, Matusop A, Ratnam S, Plasmodium knowlesi malaria in humans is widely distributed and potentially life threatening. Clin Infect Dis. 2008;46:165–71. DOIPubMedGoogle Scholar
- Jongwutiwes S, Putaporntip C, Iwasaki T, Sata T, Kanbara H. Naturally acquired Plasmodium knowlesi malaria in human, Thailand. Emerg Infect Dis. 2004;10:2211–3.PubMedGoogle Scholar
- Ng OT, Ooi EE, Lee CC, Lee PJ, Ng LC, Pei SW, Naturally acquired human Plasmodium knowlesi infection, Singapore. Emerg Infect Dis. 2008;14:814–6.PubMedGoogle Scholar
- Luchavez J, Espino F, Curameng P, Espina R, Bell D, Chiodini P, Human infections with Plasmodium knowlesi, the Philippines. Emerg Infect Dis. 2008;14:811–3.PubMedGoogle Scholar
- Knowles R, White RS. Studies in the parasitology of malaria. Indian Med Res Memoirs No. 18, Calcutta (India): Thacker, Spink, and Co.; 1930.
- Cohen JE. Heterologous immunity in human malaria. Q Rev Biol. 1973;48:467–89. DOIPubMedGoogle Scholar
- Chin W, Contacos PG, Collins WE, Jeter MH, Alpert E. Experimental mosquito-transmission of Plasmodium knowlesi to man and monkey. Am J Trop Med Hyg. 1968;17:355–8.PubMedGoogle Scholar
- Eyles DE, Coatney GR, Getz ME. Vivax-type malaria parasites of macaques transmissible to man. Science. 1960;131:1812–3. DOIPubMedGoogle Scholar
- Baimai V. Population cytogenetics of the malaria vector Anopheles leucosphyrus group. Southeast Asian J Trop Med Public Health. 1988;19:667–80.PubMedGoogle Scholar
- Brown WM, Yowell CA, Hoard A, Vander Jagt TA, Hunsaker LA, Deck LM, Comparative structural analysis and kinetic properties of lactate dehydrogenases from the four species of human malarial parasites. Biochemistry. 2004;43:6219–29. DOIPubMedGoogle Scholar
- Piper R, Lebras J, Wentworth L, Hunt-Cooke A, Houzé S, Chiodini P, Immunocapture diagnostic assays for malaria using Plasmodium lactate dehydrogenase (pLDH). Am J Trop Med Hyg. 1999;60:109–18.PubMedGoogle Scholar
- Torda AE, Procter JB, Huber T. Wurst: a protein threading server with a structural scoring function, sequence profiles and optimized substitution matrices. Nucleic Acids Res. 2004;32:W532–5. DOIPubMedGoogle Scholar
- Boyd MF, Kitchen SF. Simultaneous inoculation with Plasmodium vivax and Plasmodium falciparum. Am J Trop Med Hyg. 1937;17:855–61.
- Boyd MF, Kitchen SF, Matthews CB. Consecutive inoculations with Plasmodium vivax and Plasmodium falciparum. Am J Trop Med Hyg. 1938;18:141–50.
Figures
Cite This ArticleTable of Contents – Volume 14, Number 11—November 2008
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Thomas F. McCutchan, Laboratory of Malaria and Vector Research, National Institute of Allergy and Infectious Diseases, National Institutes of Health, 12735 Twinbrook Pkwy, Rockville, MD 20892, USA;
Top