Volume 20, Number 10—October 2014
Research
Clinical Isolates of Shiga Toxin 1a–Producing Shigella flexneri with an Epidemiological Link to Recent Travel to Hispañiola
Cite This Article
Citation for Media
Abstract
Shiga toxins (Stx) are cytotoxins involved in severe human intestinal disease. These toxins are commonly found in Shigella dysenteriae serotype 1 and Shiga-toxin–producing Escherichia coli; however, the toxin genes have been found in other Shigella species. We identified 26 Shigella flexneri serotype 2 strains isolated by public health laboratories in the United States during 2001–2013, which encode the Shiga toxin 1a gene (stx1a). These strains produced and released Stx1a as measured by cytotoxicity and neutralization assays using anti-Stx/Stx1a antiserum. The release of Stx1a into culture supernatants increased ≈100-fold after treatment with mitomycin C, suggesting that stx1a is carried by a bacteriophage. Infectious phage were found in culture supernatants and increased ≈1,000-fold with mitomycin C. Whole-genome sequencing of several isolates and PCR analyses of all strains confirmed that stx1a was carried by a lambdoid bacteriophage. Furthermore, all patients who reported foreign travel had recently been to Hispañiola, suggesting that emergence of these novel strains is associated with that region.
Shiga toxins (Stx) are potent AB5 cytotoxins that inhibit eukaryotic protein synthesis, eventually leading to host cell death (1). Infections with bacteria that produce Stx result in hemorrhagic colitis and can lead to serious complications like hemolytic uremic syndrome (HUS) (2). Although 4 species of Shigella cause bacillary dysentery, historically only Shigella dysenteriae type 1 has been recognized as carrying the genes for Shiga toxin (stx). Likewise, S. dysenteriae 1 is the only Shigella species that causes HUS as a complication of infection (3).
The genes encoding the toxin are found in an operon consisting of stxA and stxB. The stx locus in S. dysenteriae 1 is surrounded by DNA sequence homologous to lambdoid bacteriophage sequence; however, the toxin genes are not associated with a complete prophage genome (4,5). Insertion sequences flanking the stx region suggest that gene rearrangements occurred and resulted in a defective phage. As a consequence, viable phage are not recovered from S. dysenteriae 1 cultures even under conditions that induce phage production (6).
Stx has been extensively studied in Shiga toxin–producing Escherichia coli (STEC), notably E. coli O157:H7. STEC produce 2 variants of Stx: Stx1a (which differs from S. dysenteriae 1 Stx by 1 aa), and Stx2 (which shares 56% identity with Stx1a) (7,8). In contrast to the toxin genes in S. dysenteriae 1, those in STEC are generally carried by lambdoid prophages, which integrate into the host bacterial chromosome (9). The phage remains in a lysogenic state until environmental conditions induce expression of phage lytic cycle genes, leading to new phage production and lysis of the host bacterium. The stx1a and stx2 loci are found within the late gene regions of the phage; therefore, induction of the lytic cycle increases expression of the toxin genes and host cell lysis allows for toxin release (10).
Recently, acquisition of stx genes in clinical isolates of other Shigella species has been reported (11,12). Three cases of infection with S. dysenteriae 4 were described, and all were shown to express stx1. No further characterization of the stx1-encoding S. dysenteriae 4 strains was reported; however, all 3 infected patients had reported recent travel to Hispañiola (11). An isolate of Shiga toxin–producing S. sonnei from a patient returning from the Ukraine was also characterized; the toxin genes were determined to be carried by a lambdoid prophage homologous to stx-encoding phages found in STEC (13).
We identified 26 clinical isolates of S. flexneri 2 that encode stx1a. DNA sequence and PCR analyses determined that stx1a is encoded by a lambdoid prophage. Characterization of the phage indicated that it behaves similarly to stx-encoding phages that infect STEC. Like the patients from whom stx1-encoding S. dysenteriae 4 was isolated, patients from whom stx1a-encoding S. flexneri 2 was isolated and who reported foreign travel had also recently visited Hispañiola. The potential consequences of an epidemiological link to this region are discussed.
Bacterial Strains and Growth Conditions
Shigella clinical isolates used in this study are listed in Table 1. S. flexneri strains were grown in Tryptic Soy Broth (BD Difco, Franklin Lakes, NJ, USA) at 37°C with aeration or on Tryptic Soy Broth plates containing 1.5% agar and 0.025% Congo red. E. coli K-12 strain MG1655 was grown in Luria-Bertani broth and on Luria-Bertani agar plates. Kanamycin and ampicillin were used at 50 μg/mL and 100 μg/mL, respectively.
PCR Analysis of stx1a-Encoding S. flexneri
DNA lysates were used for PCR for stx1a with primers stx1-det-F1 and stx1-seq-R1, and for stx2 with primers stx_F/stxR1. All strains were verified to contain the virulence plasmid of S. flexneri by PCR amplification of virF with primers VirF1 and VirF2.
To show that stx1a was phage encoded, we analyzed lysates by PCR with primer pairs Stx1R2/Phage_stxR2 and Phage_stx1F2/Stx1F2. The insertion site of the phage into S. flexneri locus S1742 was determined by amplifying the upstream region of S1742 and an early phage gene with primers S1742_up/Stx_phage_up and by amplifying a late phage gene and the downstream region of S1742 with primers Stx_phage_dn/S1742_dn. All PCRs were conducted by using PCR Master Mix (2X) (Fermentas, Pittsburgh, PA, USA) according to the manufacturer’s specifications. Primer sequences and expected amplicons are listed in Table 2.
Construction of recA Mutants
recA was replaced with a kanamycin-resistance cassette by using λ red recombination (15). Primers RecAko-site1 and RecAko-site2 were used to amplify kan from pKD4 with 5′ and 3′ overhangs homologous to internal regions of recA. Kanamycin-resistant colonies of BS766 (15) were double purified and screened by PCR with primers RecA-1/RecA-3 for detection of the size difference between chromosomal recA and the kanamycin-resistance cassette. This mutant was used as the donor for growing a P1L4 lysate, which was used to transduce the recA::kan mutation into BS937, BS938, and BS974. Kanamycin-resistant transductants were purified and confirmed by use of PCR, as described above.
Cytotoxicity Assay
The cytotoxicity of bacterial samples for Vero cells was determined as previously described (16,17). In brief, 100 μL of diluted samples was overlain in 96-well plates containing confluent monolayers of Vero cells and incubated for 48 hours at 37°C in 5% CO2. Viable cells were fixed with 10% formalin and stained with 0.13% crystal violet. The optical density (OD) of the stained wells was measured at 630 nm by using a BioTek (Winooski, VT, USA) EL800 spectrophotometric plate reader. The CD50 (cytotoxic dose that kills 50% of the cells) was calculated by determining the inverse dilution of the bacterial sample that was required to kill 50% of the Vero cells.
In Vitro Neutralization of Stx1a
Overnight supernatants were serially diluted 10-fold in medium. We mixed 100 μL of diluted samples with 100 μL medium, a 1:25 dilution of F45 polyclonal anti-Stx1/Stx1a antiserum (17,18), or a 1:25 dilution of rabbit polyclonal antiserum against S. flexneri whole cell lysate. Toxin samples were incubated with antibody for 2 hours at 37°C in 5% CO2. We then applied 100 μL of the toxin–antibody mixture to Vero cells and incubated as above.
Mitomycin C Induction of Bacterial Lysis, Shiga Toxin 1a Production, and Prophage Induction
Overnight cultures of bacteria were inoculated 1:100 into Tryptic Soy Broth, and 2 hours after inoculation a final volume of 0.5 μg/mL mitomycin C (Sigma, St. Louis, MO, USA) was added to the cultures. To monitor the induction of bacterial lysis, we read the OD600 hourly over a period of 8 hours. Induction of bacterial lysis was noted as a 3-4–fold decrease in OD600 compared with the stx1a-negative control strains.
To determine the effect of mitomycin C on production of Stx1a and prophage induction, we collected whole cell lysates and supernatants 3 hours after addition of mitomycin C. Samples were then analyzed for cytotoxicity on Vero cells. For isolation of phage particles, supernatants were prepared similarly, except that a final concentration of 10 mmol/L MgSO4 and a drop of chloroform were added after centrifugation. As described previously, 100 μL of phage lysate was absorbed onto 200 μL of E. coli MG1655 for 20 minutes at 37°C (19). Molten L-agar top agar (19) containing 10 mmol/L MgSO4 was added to the phage/bacteria mixture and poured onto L-agar plates. Plates were incubated overnight at 37°C, and plaque-forming units (PFUs) were counted.
Isolation of Lysogens
Supernatants containing phage were prepared from mitomycin C–induced culture of BS937. Phage lysate was spotted onto an L-soft agar overlay containing either E. coli MG1655 or S. flexneri 2457T and incubated overnight at 37°C. A loop from the zone of clearing was streaked for isolation of single colonies, which were subsequently screened for stx1a genes by PCR. Positive colonies from the initial screening were double colony purified, and PCR was repeated to ensure that the colonies were positive for stx1a. MG1655 lysogens were confirmed to not be contaminates of the donor strain, BS937, by testing for S. flexneri chromosomal and virulence plasmid genes by use of PCR primer pairs IpaHF/IpaHR and VirB01/IpaFWD (Table 2), respectively. Similarly, 2457T lysogens were analyzed by PCR as described above. Two independently isolated lysogens of MG1655 and 2457T were used for further analysis.
Virulence Assays
Virulence-associated phenotypes were determined by a gentamicin protection invasion assay in HeLa cells and by plaque formation in L2 monolayers, as previously described (20,21). Both assays were conducted 3 independent times and included technical duplicates or triplicates in each individual experiment.
Whole-Genome Sequencing
DNA was isolated from overnight cultures by using a QIAGEN DNEasy Kit (Valencia, CA, USA). Samples were prepared for sequencing by using a Nextera XT DNA Sample Preparation Kit (Illumina, San Diego, CA, USA) and sequenced on an Illumina MiSeq sequencing system. The phage sequence was assembled by mapping the reads to the reference phage NC_004913.2 by using Bowtie 2 version 2.1.0 (http://sourceforge.net/projects/bowtie-bio/files/bowtie2/2.1.0/).
Nucleotide Sequence Accession Number
The complete phage sequence of ϕPOC-J13 from strain BS937 was submitted to GenBank. The sequence is available under accession no. KJ603229.
Identification and Epidemiology of stx1a-positive S. flexneri 2 Strains
BS937 and BS938 (Table 1) were acquired from the Hawaii and Massachusetts state laboratories, which had determined the isolates to be positive for stx1a by PCR. Both strains shared the same pulsed-field gel electrophoresis (PFGE) XbaI pattern, JZXX01.0357, as indicated in the Centers for Disease Control and Prevention PulseNet database (http://www.cdc.gov/pulsenet). To identify other clinical isolates of S. flexneri 2 that might be stx1a-positive, we searched the PulseNet database for strains that matched this PFGE pattern. From state public health laboratories, we obtained 18 additional strains of S. flexneri that matched this PFGE pattern. We also received time-matched, but not PFGE-matched, strains of S. flexneri as negative controls. Six additional strains that had been confirmed to be stx1a-positive S. flexneri were acquired from the Centers for Disease Control and Prevention. Clinical strains included in this study and their sources are listed in Table 1.
The stx1a-positive S. flexneri strains had been isolated over 13 years (2001–2013). They were isolated during all months except August, indicating that seasonality is not involved in the emergence and/or spread of stx1a-encoding S. flexneri. Among patients from whom stx1a-positive S. flexneri strains were isolated, no incidences of HUS were reported, suggesting that the stx1a-positive S. flexneri strains did not cause more severe disease than would typically be caused by such strains lacking stx1a. Among 22 patients for whom travel information was available, 9 reported no foreign travel or knowledge of contact with persons who had traveled. The 13 patients who reported travel had all recently visited Hispañiola or interacted with a traveler who had returned from this region (Table 1).
Verification of Stx1a in S. flexneri Isolates
The presence of stx1a in the S. flexneri strains was confirmed by PCR (data not shown). All isolates that matched PFGE pattern JZXX01.0357 or had previously been shown to encode stx1a yielded a PCR product of the correct size for the toxin gene. PCR analysis for stx2 did not produce a product. The 3 negative controls did not generate a PCR product for either stx1a or stx2.
Figure 1

Figure 1. Shiga toxin 1a (Stx1a) is produced and released from stx1a–encoding Shigella flexneri isolates. Overnight supernatants from BS937, BS938, and BS974 were serially diluted 10-fold in medium alone, medium containing anti-Stx/Stx1a antiserum,...
To determine if Stx1a was released from the bacteria, we tested supernatants from overnight cultures in a Vero cell cytotoxicity assay. All stx1a-positive isolates released a toxin into the supernatant, which killed Vero cells. To confirm that the toxin responsible for killing the cells was Stx1a, we tested overnight supernatants from 3 representative isolates for Vero cell cytotoxicity after neutralization by anti-Stx/Stx1a antiserum. After neutralization, supernatants were no longer cytotoxic to Vero cells (Figure 1). These findings demonstrate that the extracellular product responsible for cytotoxicity to Vero cells is indeed Stx1a and not a different protein being released by the stx1a-expressing S. flexneri.
Effects of Mitomycin C on Stx1a and Prophage Production
stx is generally found encoded by functional prophages (22). The prophage lytic cycle can be induced with DNA damaging agents (23); therefore, to address whether the toxin carried in stx1a-encoding S. flexneri was associated with a prophage, we tested sensitivity to lysis when grown in the presence of mitomycin C. All Stx1a-producing S. flexneri isolates showed a sharp decrease in OD600 within 3–4 hours after addition of mitomycin C, whereas the stx1a-negative strains showed no decrease in OD600 (data not shown).
Figure 2

Figure 2. Mitomycin C induces production of Shiga toxin 1a (Stx1a) in a recA-dependent manner. Exponentially growing cultures of the indicated parental strains or recA mutants were grown with or without 0.5 μg/mL...
Figure 3

Figure 3. Infectious phage in the supernatants of Shiga toxin 1a gene (stx1a)–encoding Shigella flexneri are induced with mitomycin C (mito C) treatment. Supernatants were collected from exponential cultures of BS937, BS938, and...
Because all stx1a-encoding S. flexneri isolates behaved similarly in the assays, we selected 3 isolates (BS937, BS938, and BS974) to characterize more thoroughly. To further investigate the response to mitomycin C, we grew log-phase cultures and collected samples to measure cell-associated and released toxin. Supernatants and whole cell lysates from bacteria treated with mitomycin C exhibited elevated cytotoxicity to Vero cells (Figure 2). To determine if Stx1a–producing S. flexneri generated infectious phage, we analyzed supernatants from untreated and mitomycin C–treated cultures in a PFU assay. After treatment with mitomycin C, PFUs in supernatants of BS937, BS938, and BS974 increased ≈1,000-fold (Figure 3).
Induction of the prophage lytic cycle by mitomycin C is caused by the SOS response (24). During the SOS response, the bacterial protease RecA becomes active and cleaves the phage repressor cI, which maintains the phage in a quiescent state under non-SOS conditions. Cleavage of cI enables transcription of the phage antiterminator Q, which activates the late phage genes, including stx (25). STEC recA mutants no longer release toxin or respond to agents that trigger the SOS response (24,26,27).
To investigate whether similar regulation occurs in Stx1a-producing S. flexneri, we constructed recA deletions in BS937, BS938, and BS974. recA mutants were cultured with mitomycin C as above, and samples were collected to measure the presence of toxin and phage. In the absence of mitomycin C, the recA mutants produced and released toxin in amounts comparable to those of the parental strains; however, when cultured with mitomycin C, the recA mutants did not exhibit increased cytotoxicity to Vero cells (Figure 2). Additionally, in the absence or presence of mitomycin C, no PFUs were enumerated from the recA mutants. Collectively, these data suggest that stx1a in S. flexneri is carried by a lambdoid prophage.
stx1a Carriage by a Lambdoid Prophage in S. flexneri
Figure 4
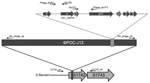
Figure 4. Schematic of PCR designed to determine that Shiga toxin 1a gene (stx1a) is phage encoded and inserted into the S1742 locus of Shigella flexneri. The genetic map shows the insertion of...
To identify the location of stx1a, we sequenced BS937, BS938, and BS974. Whole-genome sequencing confirmed that stx1a was encoded within a 62-kb lambdoid prophage. To extend the analysis to all the clinical isolates in this study, we used the PCR strategy and primer design shown in Figure 4. To ensure that stx1a in all isolates was phage encoded, we designed primers to amplify from stxA1a and stxB1a (encoding subunits A and B of Stx1a) and 1 kb either upstream or downstream of the stx1a operon. All stx1a-positive strains yielded a PCR product consistent with the toxin being phage encoded. No DNA was amplified from the stx1a-negative isolates.
Figure 5

Figure 5. PCR results from representative clinical isolates illustrate that the Shiga toxin 1a gene (stx1a) is phage encoded and inserted into the S1742 locus of Shigella flexneri. PCRs based on the primer...
Whole-genome sequencing revealed that the phage was inserted into locus S1742 (which encodes a putative oxidoreductase) of the S. flexneri chromosome. Primers were designed (Figure 4) to determine if the stx1a-encoding phage inserted into S1742 for all isolates. All stx1a-positive strains generated the expected PCR product when amplified with primers specific for the early phage sequence and upstream of S1742 and with primers directed to the late phage sequence and downstream of S1742. None of the stx1a-negative strains yielded an amplified product with the primer pairs. A representative gel of the 4 amplifications for 1 stx1a-negative and 6 stx1a-positive isolates is shown in Figure 5. These data suggest that a phage has integrated into all stx1a-positive isolates. We named this stx1a-encoding phage ϕPOC-J13.
Lysogeny of Laboratory Strains of E. coli and S. flexneri with ϕPOC-J13
E. coli MG1655 and S. flexneri 2457T were lysogenized as described earlier. To test for production and release of Stx1a, we examined whole cell lysates and supernatants from overnight cultures of MG1655 and 2457T lysogens in Vero cell cytotoxicity assays. The average CD50/mL of supernatants from MG1655 and 2457T lysogens was 1 ×105, similar to that of the stx1a-positive S. flexneri clinical isolates. Lysogens were also tested for the presence of phage in supernatants of overnight cultures by determining PFUs. The 2457T lysogens released ≈106 PFUs/mL; however, no viable phage could be recovered from MG1655 lysogens.
To confirm that stx1a in the lysogens was phage associated, we conducted PCR amplification as described above (Figure 4). The MG1655 and 2457T lysogens yielded a PCR product that indicated that the phage regions upstream and downstream of stx1a were present. Lysogens were also tested for integration of the phage into locus S1742 by use of the primers illustrated in Figure 4. In 2457T lysogens, ϕPOC-J13 inserted into S1742 and produced a PCR product of the expected size. Similarly, in lysogens of MG1655, ϕPOC-J13 inserted into ynfG, the E. coli S1742 homologue. These findings demonstrate that the integration of ϕPOC-J13 is site specific, as has been shown for other lambdoid prophages (9).
Virulence Phenotypes of Shiga Toxin 1a–producing S. flexneri
We wanted to determine if the presence of ϕPOC-J13 altered the virulence phenotypes associated with S. flexneri. Invasion and plaquing efficiencies of BS937, BS938, and BS974 were compared with those of laboratory strain 2457T (Table 3). BS938 and BS974 exhibited similar invasion efficiency as 2457T; however, invasion with BS937 was significantly higher. All 3 Stx1a–producing S. flexneri isolates showed plaquing efficiency comparable to that of 2457T, and the plaque diameters were consistent among all strains. Although it is unclear why BS937 was more invasive, the comparable level of cell-to-cell spread suggests that ϕPOC-J13 does not appreciably alter the virulence properties of S. flexneri.
Bacteriophages are recognized for their contribution to the genetic diversity of bacteria and for their capacity to transfer virulence factors (28). It was first noted in the early 1980s that stx in E. coli was encoded by a lambdoid bacteriophage (29,30). We have identified a new stx1a-encoding bacteriophage, ϕPOC-J13, from clinical isolates of S. flexneri. Generally, the acquisition of toxin genes is thought to increase the virulence of a bacterial species. However, according to the available clinical data and our in vitro virulence assays, the production of Stx1a in S. flexneri does not seem to increase pathogenicity within the host.
Characterization of ϕPOC-J13 determined that it behaves similarly to stx-encoding phages found in STEC; however, some differences are notable. First, although ϕPOC-J13 responded to DNA damaging treatment by inducing the lytic cycle and induction was RecA-dependent, recA mutants of S. flexneri Stx1a–producing strains still maintained a level of Stx1a production and release comparable to that of the noninduced parental strains. In contrast, recA mutants of STEC produce a very low level of Stx, and the toxin that is present remains largely cell associated rather than being released into the supernatant (24,26,27). One explanation for the differences in phenotype between Stx1a-producing S. flexneri recA mutants and the STEC mutants might be that all STEC recA mutants examined encoded stx2; thus, the regulation of stx1a- and stx2-encoding phages might vary. Additionally, although Stx1a was produced in S. flexneri recA mutants, viable phage particles were not recovered. stx1a has an upstream promoter that is not dependent on induction of the phage lytic cycle (31). A similar promoter might be responsible for the baseline level of Stx1a produced in the S. flexneri recA mutants and would explain the lack of infectious phage in the mutants.
ϕPOC-J13 lysogenized laboratory strains of E. coli and S. flexneri. Viable phage particles were recovered from the supernatants of 2457T lysogens but not from those of MG1655 lysogens, even though Stx1a was produced and released by lysogens of both species. This result might suggest that the stability and/or assembly of ϕPOC-J13 varies according to the host bacterium. Host differences in the regulation of ϕPOC-J13 might also account for the discrepancies between the recA mutants of Stx1a-producing S. flexneri and STEC. Our future studies will compare the differences in regulation of stx1a in ϕPOC-J13 with that of known STEC phages.
Another aspect of these stx1a-encoding S. flexneri isolates is their potential epidemiological link to Hispañiola. Although some patients reported no travel, ≈60% reported travel to this region or interaction with a traveler returning from this region. Most of our clinical isolates came from public health laboratories in the eastern United States, suggesting a possible focus in that area. However, the large number of isolates from the eastern United States might simply reflect the large Haitian immigrant population in this region and the resultant frequent travel to Haiti (32). Nevertheless, further surveillance of Stx1a–producing S. flexneri is warranted to determine the extent of their emergence in Hispañiola.
The epidemiological link to Hispañiola generates many questions about what has led to the emergence of these strains. The earliest Stx1a–producing S. flexneri isolates pre-date the earthquake that struck Haiti in January 2010. Thus, this natural disaster is not linked to the presence of stx1a-encoding S. flexneri in the region. One possibility is that the ecosystem in Hispañiola is favorable for the acquisition of ϕPOC-J13 by S. flexneri and possibly other Shigella species. An environmental reservoir of Shigella spp. has never been identified; therefore, it is tempting to speculate that production of Stx1a might give S. flexneri a survival advantage in the aquatic environment. In accordance with this hypothesis, studies on the survival of Shigella spp. in amebae indicate that S. dysenteriae 1 can persist longer than S. flexneri within Acanthamoeba castellanii (33,34). In addition, Stx–producing bacteria can kill the protozoan Tetrahymena thermophile to avoid consumption by this predator (35). Thus, Stx1a might benefit S. flexneri by providing a defense against eukaryotic predators.
It will be important to study clinical isolates of other Shigella species and bacterial genera to determine whether they also harbor ϕPOC-J13; we expect that the occurrence of this stx1a-encoding phage will be more widespread. Although toxin production in S. flexneri did not suggest an increase in pathogenicity, the consequences of the emergence of such Stx1a–producing strains are impossible to predict. Future studies that address these questions will provide a better understanding of the emergence of stx1a-encoding S. flexneri.
Dr Gray is a postdoctoral research fellow at Uniformed Services University of the Health Sciences. Her primary research interests focus on understanding bacterial pathogens that cause intestinal diseases.
Acknowledgments
We thank the state health laboratories that provided strains, Charlie Wang and James Pettengill who provided sequencing assistance, and Stephen Darnell who provided technical assistance.
This work was supported by grants R01AI24656 and AI020148 from the National Institute of Allergy and Infectious Diseases.
References
- Tesh VL, O'Brien AD. The pathogenic mechanisms of Shiga toxin and the Shiga-like toxins. Mol Microbiol. 1991;5:1817–22 . DOIPubMedGoogle Scholar
- Butler T. Haemolytic uraemic syndrome during shigellosis. Trans R Soc Trop Med Hyg. 2012;106:395–9. DOIPubMedGoogle Scholar
- Keusch GT, Bennish ML. Shigellosis: recent progress, persisting problems and research issues. Pediatr Infect Dis J. 1989;8:713–9. DOIPubMedGoogle Scholar
- McDonough MA, Butterton JR. Spontaneous tandem amplification and deletion of the Shiga toxin operon in Shigella dysenteriae 1. Mol Microbiol. 1999;34:1058–69. DOIPubMedGoogle Scholar
- Unkmeir A, Schmidt H. Structural analysis of phage-borne stx genes and their flanking sequences in Shiga toxin–producing Escherichia coli and Shigella dysenteriae type 1 strains. Infect Immun. 2000;68:4856–64. DOIPubMedGoogle Scholar
- Strockbine NA, Jackson MP, Sung LM, Holmes RK, O'Brien AD. Cloning and sequencing of the genes for Shiga toxin from Shigella dysenteriae type 1. J Bacteriol. 1988;170:1116–22 .PubMedGoogle Scholar
- Calderwood SB, Auclair F, Donohue-Rolfe A, Keusch GT, Mekalanos JJ. Nucleotide sequence of the Shiga-like toxin genes of Escherichia coli. Proc Natl Acad Sci U S A. 1987;84:4364–8. DOIPubMedGoogle Scholar
- Newland JW, Strockbine NA, Neill RJ. Cloning of genes for production of Escherichia coli Shiga-like toxin type II. Infect Immun. 1987;55:2675–80 .PubMedGoogle Scholar
- Schmidt H. Shiga-toxin-converting bacteriophages. Res Microbiol. 2001;152:687–95. DOIPubMedGoogle Scholar
- Neely MN, Friedman DI. Functional and genetic analysis of regulatory regions of coliphage H-19B: location of Shiga-like toxin and lysis genes suggest a role for phage functions in toxin release. Mol Microbiol. 1998;28:1255–67. DOIPubMedGoogle Scholar
- Gupta SK, Strockbine N, Omondi M, Hise K, Fair MA, Mintz E. Emergence of Shiga toxin 1 genes within Shigella dysenteriae type 4 isolates from travelers returning from the Island of Hispanola. Am J Trop Med Hyg. 2007;76:1163–5 .PubMedGoogle Scholar
- Beutin L, Strauch E, Fischer I. Isolation of Shigella sonnei lysogenic for a bacteriophage encoding gene for production of Shiga toxin. Lancet. 1999;353:1498. DOIPubMedGoogle Scholar
- Strauch E, Lurz R, Beutin L. Characterization of a Shiga toxin–encoding temperate bacteriophage of Shigella sonnei. Infect Immun. 2001;69:7588–95. DOIPubMedGoogle Scholar
- Scheutz F, Teel LD, Beutin L, Pierard D, Buvens G, Karch H, Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J Clin Microbiol. 2012;50:2951–63. DOIPubMedGoogle Scholar
- Clark CS, Maurelli AT. Shigella flexneri inhibits staurosporine-induced apoptosis in epithelial cells. Infect Immun. 2007;75:2531–9. DOIPubMedGoogle Scholar
- Gentry MK, Dalrymple JM. Quantitative microtiter cytotoxicity assay for Shigella toxin. J Clin Microbiol. 1980;12:361–6 .PubMedGoogle Scholar
- Schmitt CK, McKee ML, O'Brien AD. Two copies of Shiga-like toxin II–related genes common in enterohemorrhagic Escherichia coli strains are responsible for the antigenic heterogeneity of the O157:H- strain E32511. Infect Immun. 1991;59:1065–73 .PubMedGoogle Scholar
- O’Brien AD, LaVeck GD. Purification and characterization of a Shigella dysenteriae 1–like toxin produced by Escherichia coli. Infect Immun. 1983;40:675–83 .PubMedGoogle Scholar
- Lech K, Brent R. Plating lambda phage to generate plaques. Curr Protoc Mol Biol. 2001;Chapter 1:Unit1.11. PMID: 18265039 .DOIGoogle Scholar
- Kane CD, Schuch R, Day WA Jr, Maurelli AT. mxiE regulates intracellular expression of factors secreted by the Shigella flexneri 2a type III secretion system. J Bacteriol. 2002;184:4409–19. DOIPubMedGoogle Scholar
- Zurawski DV, Mitsuhata C, Mumy KL, McCormick BA, Maurelli AT. OspF and OspC1 are Shigella flexneri type III secretion system effectors that are required for postinvasion aspects of virulence. Infect Immun. 2006;74:5964–76 . DOIPubMedGoogle Scholar
- Herold S, Karch H, Schmidt H. Shiga toxin–encoding bacteriophages—genomes in motion. Int J Med Microbiol. 2004;294:115–21 . DOIPubMedGoogle Scholar
- Köhler B, Karch H, Schmidt H. Antibacterials that are used as growth promoters in animal husbandry can affect the release of Shiga-toxin-2–converting bacteriophages and Shiga toxin 2 from Escherichia coli strains. Microbiology. 2000;146:1085–90 .PubMedGoogle Scholar
- Fuchs S, Muhldorfer I, Donohue-Rolfe A, Kerenyi M, Emody L, Alexiev R, Influence of RecA on in vivo virulence and Shiga toxin 2 production in Escherichia coli pathogens. Microb Pathog. 1999;27:13–23. DOIPubMedGoogle Scholar
- Pacheco AR, Sperandio V. Shiga toxin in enterohemorrhagic E.coli: regulation and novel anti-virulence strategies. Front Cell Infect Microbiol. 2012;2:81.
- Mühldorfer I, Hacker J, Keusch GT, Acheson DW, Tschape H, Kane AV, Regulation of the Shiga-like toxin II operon in Escherichia coli. Infect Immun. 1996;64:495–502 .PubMedGoogle Scholar
- Muniesa M, Recktenwald J, Bielaszewska M, Karch H, Schmidt H. Characterization of a Shiga toxin 2e-converting bacteriophage from an Escherichia coli strain of human origin. Infect Immun. 2000;68:4850–5. DOIPubMedGoogle Scholar
- Brüssow H, Canchaya C, Hardt WD. Phages and the evolution of bacterial pathogens: from genomic rearrangements to lysogenic conversion. Microbiol Mol Biol Rev. 2004;68:560–602 . DOIPubMedGoogle Scholar
- O'Brien AD, Newland JW, Miller SF, Holmes RK, Smith HW, Formal SB. Shiga-like toxin-converting phages from Escherichia coli strains that cause hemorrhagic colitis or infantile diarrhea. Science. 1984;226:694–6. DOIPubMedGoogle Scholar
- Scotland SM, Smith HR, Willshaw GA, Rowe B. Vero cytotoxin production in strain of Escherichia coli is determined by genes carried on bacteriophage. Lancet. 1983;322:216. DOIPubMedGoogle Scholar
- Wagner PL, Livny J, Neely MN, Acheson DW, Friedman DI, Waldor MK. Bacteriophage control of Shiga toxin 1 production and release by Escherichia coli. Mol Microbiol. 2002;44:957–70 . DOIPubMedGoogle Scholar
- Louis BM. Geographies of the Haitian diaspora. Transforming Anthropol. 2013;21:198–9. DOIGoogle Scholar
- Saeed A, Abd H, Edvinsson B, Sandstrom G. Acanthamoeba castellanii an environmental host for Shigella dysenteriae and Shigella sonnei. Arch Microbiol. 2009;191:83–8. DOIPubMedGoogle Scholar
- Saeed A, Johansson D, Sandstrom G, Abd H. Temperature depended role of Shigella flexneri invasion plasmid on the interaction with Acanthamoeba castellanii. Int J Microbiol. 2012;•••:1–8. DOIPubMedGoogle Scholar
- Lainhart W, Stolfa G, Koudelka GB. Shiga toxin as a bacterial defense against a eukaryotic predator, Tetrahymena thermophila. J Bacteriol. 2009;191:5116–22. DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 20, Number 10—October 2014
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Anthony T. Maurelli, Department of Microbiology and Immunology, F. Edward Hébert School of Medicine, Uniformed Services University of the Health Sciences, 4301 Jones Bridge Rd, Bethesda, MD 20814-4799, USA
Top