Volume 24, Number 8—August 2018
Research
Human Norovirus Replication in Human Intestinal Enteroids as Model to Evaluate Virus Inactivation
Cite This Article
Citation for Media
Abstract
Human noroviruses are a leading cause of epidemic and endemic acute gastroenteritis worldwide and a leading cause of foodborne illness in the United States. Recently, human intestinal enteroids (HIEs) derived from human small intestinal tissue have been shown to support human norovirus replication. We implemented the HIE system in our laboratory and tested the effect of chlorine and alcohols on human norovirus infectivity. Successful replication was observed for 6 norovirus GII genotypes and was dependent on viral load and genotype of the inoculum. GII.4 viruses had higher replication levels than other genotypes. Regardless of concentration or exposure time, alcohols slightly reduced, but did not completely inactivate, human norovirus. In contrast, complete inactivation of the 3 GII.4 viruses occurred at concentrations as low as 50 ppm of chlorine. Taken together, our data confirm the successful replication of human noroviruses in HIEs and their utility as tools to study norovirus inactivation strategies.
Human noroviruses are the leading cause of epidemic and endemic acute gastroenteritis worldwide (1). A major barrier to studying the pathogenesis, virus–host interactions, and effect of control measures to prevent and treat norovirus gastroenteritis has been the lack of a robust and reproducible cell culture system. Since the discovery of norovirus in 1972, many research groups have attempted to grow human norovirus. Initial studies included a comprehensive number of primary and continuous cell lines that had been used successfully to grow other viruses, but none supported human norovirus replication (2). Other attempts included the use of a differentiated human embryonic small-intestinal cell line (INT 407) (3), but the results could not be confirmed by others (4–6). The discovery of murine norovirus (MNV) in 2003 and the fact that this virus successfully replicated in a murine macrophage cell line in vitro and in primary immune cells in vivo suggested that immune cells may also support replication of human norovirus (7). However, immune cells isolated from healthy adults did not support replication (8). A more recent study reported that BJAB cells, a continuous human B cell line, supported replication of a GII.4 norovirus strain in the presence of bacteria (9). Although initially promising, these results have not been consistently confirmed by other groups (10).
The first steps toward a new cell culture system for human norovirus included the detection of human norovirus antigen in duodenal and jejunal enterocytes in tissue sections from human norovirus–infected transplant patients (11) and the development of human intestinal enteroids (HIEs) derived from nontransformed small intestine and colonic tissues (12). The HIEs, or “mini-guts,” are generated from stem cells present in the intestinal crypts isolated from human intestinal tissue and cultured indefinitely as ex vivo, 3-dimensional (3D) cultures in growth-factor–enriched media (13–15). HIE cultures recapitulate the complexity and cell diversity of the gastrointestinal tract in relatively the same proportions as in the intestine itself (14,15) and successfully support the replication of human rotavirus (13) and human norovirus (16,17). These studies confirmed the role of enterocytes as the major site for human norovirus replication and host restriction based on genetic factors, as well as the role of bile as a strain-specific requirement or enhancer for virus infectivity.
During the past 40 years, the efficacy of inactivation and disinfection procedures for human norovirus could be evaluated only by human challenge studies (18,19) or by using cultivable surrogate viruses such as MNV, feline calicivirus, or Tulane virus (20–23). However, without direct confirmation that any of these surrogates correlate with inactivation of infectious human norovirus, no consensus has been reached on the best surrogate for human norovirus (24,25). In this study, we demonstrate the successful implementation of HIE cultures, show successful replication of several human norovirus genotypes, and demonstrate the applicability of HIEs to evaluate the efficacy of chlorine and alcohols on reducing virus infectivity.
Detailed methods and description of HIE cultures, gene expression analysis, viral infections, norovirus detection, and statistical analyses are provided in the Technical Appendix. This investigation was determined by the Centers for Disease Control and Prevention (CDC) to be public health nonresearch and therefore not subject to institutional review board review.
Fecal Samples
We included 80 human norovirus-positive fecal samples (12 genogroup [G] I, 65 GII, and 3 GIV) collected during 2000–2017 in the study (Table 1). Samples were stored at 4°C or −70°C from collection time until the time of testing. All samples were tested during April 2016–December 2017.
Human Intestinal Enteroid Culture
Baylor College of Medicine (Houston, TX, USA) provided secretor-positive jejunal HIE cultures (J2 and J3 lines) and Wnt3a-producing cells (CRL-2647 cells). Calvin Kuo (Palo Alto, CA, USA) kindly provided R-spondin-producing cells. Gijs van den Brink (University of Amsterdam, Amsterdam, the Netherlands) kindly provided Noggin-producing cells. Complete media with growth factors (CMGF+) and without growth factors (CMGF–), differentiation media, and 3 conditioned media (Wnt3a, R-spondin, and Noggin) were prepared as reported previously (13,16,17).
We grew jejunal HIE cultures (J2 or J3 lines) as undifferentiated 3D cultures in the presence of CMGF+ supplemented with 10 μmol/L Y-27632, as described previously (17). After 7 days, highly dense 3D cultures were either split 1:2, frozen in LN2, or dissociated into a single cell suspension and plated as undifferentiated monolayers. After culture for 24 h at 37°C in 5% CO2, we replaced CMGF+ supplemented with 10 μmol/L Y-27632 with differentiation medium to induce monolayer differentiation.
Infection Experiments and Viral Replication
We performed all infections in triplicate on 100% confluent 4-day-old differentiated HIE (J3 line) monolayers, except when specified that the J2 line was used. In some experiments, we pretreated monolayers with 1% sow bile included in the differentiation medium 48 h before infection. In other experiments, we differentiated HIE monolayers without pretreatment, and infected them in the presence of 500 μmol/L of glycochenodeoxycholic acid (GCDCA; Sigma, St. Louis, MO, USA) or with 500 μM GCDCA plus 50 μM ceramide.
To determine viral infectivity, we inoculated duplicate 96-well plates with 100 μL of fecal filtrate (Technical Appendix) at 1:10, 1:100, and 1:1000 dilution. After 1 h incubation at 37°C in 5% CO2, we washed the monolayers twice with CMGF– and added 100 μL of differentiation medium containing 1% sow bile, 500 μmol/L GCDCA, or 500 μM GCDCA plus 50 μmol/L ceramide to each well. For each set of infections, we immediately froze 1 plate at −70°C and incubated a duplicate plate at 37°C in 5% CO2 for 72 h and froze it at −70°C. We used quantitative reverse transcription real-time PCR to determine the amount of norovirus RNA from input virus and from HIE monolayers at 1 hour postinfection (hpi) and at 72 hpi. Standard curve based on quantified RNA transcripts was included.
Inactivation Treatments
Alcohol Treatment
We diluted 10% fecal filtrates 1:10 in 70% ethanol or isopropanol solutions and incubated them for 1 min or 5 min. We then neutralized the alcohols in the samples by adding 9 volumes of CMGF supplemented with 10% fetal bovine serum (FBS). We included a nontreatment control and an alcohol neutralization control in each experiment.
Chlorine Suspension Assays
We prepared fresh chlorine stock solutions at 1,000 ppm and 10,000 ppm by diluting commercial bleach (6% sodium hypochlorite) in cell culture–grade water. We diluted 20 μL of 10% fecal filtrates in an appropriate volume of chlorine stock solutions to achieve a series of total chlorine concentrations of 5–5000 ppm. After incubating the solutions for 1 min at room temperature, we added sodium thiosulfate (final concentration 50 mg/L) to neutralize free chlorine. We included a nontreatment control and a chlorine neutralization control in each experiment.
HIE Model and Small Intestine Complexity
Figure 1
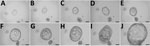
Figure 1. Growth of 3-dimensional human intestinal enteroid (HIE) embedded in Matrigel in the presence of complete media with growth factors containing Wnt3a, R-spondin, and Noggin as part of a study of human...
Figure 2
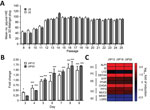
Figure 2. Characterization of differentiated and undifferentiated HIE in a study of human norovirus replication in HIEs. A) Quantification of undifferentiated HIE generated on each passage. Undifferentiated HIEs derived from 2 donors (J2P7...
We recovered jejunal HIE cultures from 2 donors (J2 and J3) frozen at passage 7 (P7) from LN2 and grew them as 3D cultures in Matrigel (BD Biosciences, San Jose, CA, USA). Within 24 hours, cells formed small cystic or multilobular HIEs and continued to grow in the presence of CMGF+ medium (Figure 1). With each passage, the number of 3D HIEs doubled, reaching a maximum Matrigel capacity of 100/plug. We were able to culture HIEs for >4 months (16−17 consecutive passages) (Figure 2, panel A). We confirmed that the number of highly proliferative stem cells increased over time, as shown by enhanced transcriptional levels of LGR5+ (Figure 2, panel B), whereas differentiated monolayers’ LGR5+ expression levels were greatly reduced and lineage-specific markers (SI, ALPI, TFF3, MUC2, FFA4R) were increased (Figure 2, panel C).
Human Norovirus Infection of Jejunal HIEs
Figure 3
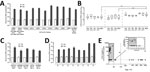
Figure 3. Evaluation of human norovirus replication in human intestinal enteroids (HIEs). A) Jejunal HIE monolayers (J3 line) inoculated with GII.4 P4 Den Haag-GII.4 Den Haag (6.2 × 104 RNA copies/well), GII.4 P4...
A previous study demonstrated the replication of GII.4 norovirus in HIEs (16). To evaluate whether those results could be reproduced, we infected jejunal HIEs (line J3) with GII.4 fecal filtrates (GII.P4 Den Haag-GII.4 Den Haag, GII.P4 New Orleans-GII.4 New Orleans, GII.Pe-GII.4 Sydney, and GII.P16-GII.4 Sydney). At 72 hpi, we detected 100- to 1,100-fold increases in viral RNA copies per well for all GII.4 fecal filtrates compared with viral RNA levels detected at 1 hpi (Figure 3, panel A; Table 2).
To further evaluate the reproducibility of the system, we included GII.4 fecal filtrates (GII.4 Den Haag, GII.4 New Orleans, and GII.4 Sydney) in each infection experiment conducted during August 2016–2017. We observed consistent replication of the 3 strains without significant differences in viral titers (Figure 3, panel B). The mean log10 increase was 2.7 (95% CI 2.68–2.82; n = 35) for GII.4 Sydney, 2.4 (95% CI 2.29–2.45; n = 33) for GII.4 Den Haag, and 2.3 (95% CI 2.17–2.46; n = 18) for GII.4 New Orleans. We observed significantly higher-fold increases on viral RNA titers for GII.4 Sydney compared with GII.4 Den Haag and GII.4 New Orleans (p<0.0001).
To evaluate whether HIEs support replication of non-GII.4 strains, we inoculated monolayers with different GI-, GII-, and GIV-positive fecal filtrates (Table 1). We observed viral replication for GII.Pg-GII.1 (2.6 log10), GII.P16-GII.2 (2.8 log10), GII.P12-GII.3 (2.3 log10), GII.P7-GII.14 (1.7 log10), and GII.Pe-GII.17 (1.9 log10) strains (Table 2; Figure 3, panel C). We did not observe replication of GI, GIV, and other GII genotypes. We also confirmed that both J2 and J3 HIE lines support human norovirus replication without significant differences in -fold change between the 2 cell lines (Figure 3, panel D). Most (15/16; 94%) samples that successfully replicated had been stored at −70°C (Table 2) and were collected from children <2 years of age (13/16; 81%) (Figure 3, panel E). CDC830 was stored at 4°C for 2 months before cultivation; this sample was collected from an adult 83 years of age.
Figure 4
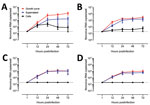
Figure 4. Evaluation as human norovirus replication in human intestinal enteroids (HIEs) by assessment of kinetics of infection for 4 GII genotypes. We inoculated jejunal HIE monolayers (J3 line) with A) GII.4 Pe-GII.4...
We next evaluated replication of human norovirus in HIEs by assessing the kinetics of infection for 4 GII genotypes (GII.1, GII.2, GII.3, and GII.4 Sydney). Consistent with a successful infection, norovirus RNA levels increased at 12 hpi, reaching a plateau at 24 hpi; no significant further increase at 72 hpi was observed for any of the genotypes (Figure 4). Despite a similar viral input level (3.3–9.3 × 105 copies/well), GII.4 Sydney infected HIEs with higher efficiency than did the other 3 genotypes (Figure 4), as shown by higher levels of viral RNA in cells and supernatant.
Figure 5

Figure 5. Confirmatory testing for human norovirus replication in human intestinal enteroids (HIEs). We inoculated jejunal HIE monolayers (J3 line) with A) GII.4 Pe-GII.4 Sydney (3.3 × 105 RNA copies/well), B) GII.P12-GII.3 (5.3...
To further confirm the production and release of norovirus from cells, we quantified viral titers in supernatants collected from cell cultures and replaced the differentiation media every 24 hours after infection (Figure 5). We detected viral RNA in supernatants collected at 24 hpi and 48 hpi for all infections. Higher and more consistent levels of norovirus RNA were detected in HIE infected with GII.4 Sydney up to 96 hpi, whereas the initial RNA levels detected for GII.1, GII.2, and GII.3 declined and became undetectable after 48 hpi for GII.1 and GII.2. These data clearly demonstrate that, although HIEs are permissive to infection with different norovirus genotypes, GII.3 and GII.4 replicated with higher efficiency.
Figure 6
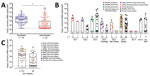
Figure 6. Comparison of amount of input viral RNA with success of human norovirus replication in human intestinal enteroids (HIEs). A) We infected HIE monolayers with undiluted or prediluted (1:10; 1:100; 1:1000) 10%...
Figure 7
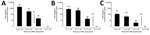
Figure 7. Determination of ID50 required for human norovirus replication in human intestinal enteroids (HIEs). We inoculated HIE monolayers in triplicate with 10-fold serial diluted fecal filtrates A) A5413_GII.4 Sydney, B) G3868_GII.4 Den...
We also compared the amount of input viral RNA with the success of replication. Samples that replicated successfully had a significantly higher input titer compared with strains that did not replicate (p<0.0001) (Figure 6, panel A). Stratified by genotype, the effect of the initial input amount of virus was observed for infections performed with GII.1, GII.2, GII.4 Den Haag, GII.4 New Orleans, and GII.4 Sydney viruses (Figure 6, panels B, C). To further confirm the role of the amount of virus inoculum on the success of replication, we infected HIE monolayers with 10-fold serial dilutions of GII.3 and GII.4 fecal filtrates. The dose required to produce infection in 50% of the inoculated wells (ID50) was 2.1 × 103 genome copies/well for GII.4 Den Haag, 4.4 × 102 genome copies/well for GII.4 Sydney, and 4.0 × 103 genome copies/well for GII.3, based on the Reed-Muench method (Figure 7) (28).
Inactivation of Human Norovirus by Alcohols
Figure 8
![Thumbnail of Inactivation of human norovirus by chlorine compared with 70% alcohol solutions. A) Ten percent fecal filtrates (G3868 [GII.4 Den Haag], 2.04 × 106 RNA copies; G3829 [GII.4 New Orleans], 4.14 × 106 RNA copies; A5413 [GII.4 Sydney], 1.58 × 107 RNA copies) were either treated or not treated with 70% ethanol for 1 min or 5 min at room temperature. We added complete media without growth factors supplemented with 10% fetal bovine serum to neutralize remaining ethanol. B) Ten percent feca](/eid/images/18-0126-F8-tn.jpg)
Figure 8. Inactivation of human norovirus with 70% alcohol or chlorine solution in suspension. A) Ten percent fecal filtrates (G3868 [GII.4 Den Haag], 2.04 × 106 RNA copies; G3829 [GII.4 New Orleans], 4.14...
We next evaluated the efficacy of alcohols to inactivate infectious human norovirus by using 3 successfully replicating GII.4 viruses (GII.4 Den Haag, GII.4 New Orleans, and GII.4 Sydney). Although replication levels of fecal filtrates exposed to 70% ethanol for 1 and 5 minutes were significantly lower compared with nontreated fecal filtrates (p<0.05), none of the GII.4 viruses was completely inactivated by ethanol (Figure 8, panel A). In addition, we treated 2 GII.4 Sydney fecal filtrates (R3702 and CDC830) with 70% ethanol or 70% isopropanol for 5 minutes to rule out the possibility that the observed inactivation patterns were sample specific (Figure 8, panel B). We observed no complete inactivation for any of the tested samples, although the replication levels after treatment with 70% ethanol were up to 0.7 log10 lower, and we found no reduction after exposure to isopropanol (Figure 8, panel B). Following treatment of the fecal filtrates with 70% isopropanol for 5 minutes, norovirus input RNA was still detectable, whereas input titers after 70% alcohol treatment were reduced (1.3 to 2.9 log10).
Inactivation of Human Norovirus by Chlorine
To evaluate the ability of chlorine to effectively inactivate infectious human norovirus, we treated fecal filtrates of 3 GII.4 viruses with increasing concentrations of chlorine (5–5,000 ppm) for 1 minute. Compared with nontreated controls, all chlorine concentrations >50 ppm completely inactivated GII.4 Den Haag, GII.4 Sydney (Figure 8, panels C, D), and GII.4 New Orleans (data not shown). Norovirus input RNA was detectable in all samples that were treated with <600 ppm of chlorine.
Since the discovery of Norwalk virus, many attempts have been made to culture human noroviruses; most efforts were unsuccessful, or the results were not reproducible in other laboratories (2–6,9,10). The successful long-term expansion of intestinal epithelial organoids has been a major breakthrough in the field of in vitro culture of intestinal epithelium (12,14,15). Recent studies show that HIEs support replication of human norovirus and other enteric viruses (13,16,29) and enable analysis of the innate immune response against these viruses (30). In this study, we showed successful replication of 6 GII norovirus genotypes (GII.1, GII.2, GII.3, GII.4, GII.14, and GII.17), including 3 GII.4 variants. Repeated infections conducted over a 1-year period showed consistent increase in viral titers of these 3 GII.4 variant strains, demonstrating that the HIE model is robust. Our data also demonstrate that, after initial confirmation of infectivity, storage of fecal samples at −70°C will preserve virus infectivity for at least 1 year.
We showed successful replication for 6 of the 14 genotypes tested in this study, although the success rate varied. Strain-specific differences have been reported for other viruses grown in HIEs (13,29). For example, compared with echovirus 11 and coxsackievirus B, enterovirus 71 replicates to significantly lower levels in HIEs (29). Enteroids also support robust replication of human rotavirus strains Ito (G3P[8]) and Wa (G1P[8]) but not the attenuated G1P[8] human rotavirus vaccine strain (13). Ettayebi et al. also demonstrated that GII.4 Sydney strains infect enteroids with higher efficiency than do GI.1, GII.3, and GII.17 viruses (16); in our study, GII.4 norovirus strains replicated at higher efficiency than did GII.1, GII.2, and GII.3 viruses. In addition, the ID50 values for GII.4 and GII.3 viruses were similar but slightly lower (2–5×) than reported previously (16). Taken together, these results indicate that high viral RNA titers are not a guarantee for suc-cessful replication, perhaps suggesting that fecal specimens that do show norovirus replication may contain large num-bers of noninfectious particles.
Until now, evaluation of control measures for human norovirus, including disinfection measures, has relied primarily on the use of cultivable surrogate viruses (24). Although these viruses are similar in size and genome organization, none completely mimics the inactivation patterns of human norovirus based on reduction of viral RNA titers. We demonstrated that the HIE model can be used to evaluate the effectiveness of alcohols and chlorine against human norovirus. Although 5 minutes of exposure to 70% ethanol and isopropanol slightly reduced viral RNA levels, overall, the alcohols did not inactivate GII.4 viruses. These results are in agreement with a previous study that, based on lack of reduction of viral RNA titers, suggested that GII human noroviruses are not affected by alcohol (24). In a comprehensive study comparing different cultivable surrogate viruses for human norovirus, Cromeans et al. (24) showed that Tulane virus, but not feline calcivirus or MNV, was resistant to alcohols. Using HIEs, we now demonstrate that human norovirus closely resembles Tulane virus when measuring inactivation by alcohols.
For chlorine, our data showed that complete inactivation of 3 different GII.4 strains could be achieved with concentrations as low as 50 ppm. These results are consistent with a recent report indicating that treatment with chlorine concentrations <50 ppm were not sufficient to inactivate human norovirus in secondary effluents from water treatment plants (31). In conclusion, our inactivation data demonstrate that chlorine, but not alcohol, completely inactivates human norovirus and that evaluation of inactivation strategies based only on detection of viral RNA does not always reflect the effectiveness of the treatment.
Our study has several limitations. First, the success rate of samples with a moderate viral RNA titer was relatively low, and thus far we have had no success with GI and GIV samples. However, because we were also not able to replicate several high viral load GII samples, other, currently unknown, factors also contribute to successful replication. Second, although we demonstrated that infectious particles are produced and we were able to measure complete inactivation by chlorine treatment, we analyzed only viral RNA levels, not protein levels. Further work is needed on the amount of chlorine required to inactivate human norovirus because we used fecal samples, which inherently have a high chlorine burden, and we measured total chlorine, whereas the level of free chlorine is what actually determines inactivation. Finally, the HIE model is costly and labor intensive; additional improvements are required to make it more affordable and widely available.
In conclusion, we confirmed that the HIE system to culture human norovirus (16) can be successfully implemented in another laboratory. The culture system supported identical levels of replication of a panel of human norovirus strains consistently for >1 year. The success of replication depends on genotype and initial virus titer but also on other unknown factors. Additional HIE cell lines need to be tested or cultures need to be enriched for enterocytes (32) because replication of some noroviruses is restricted by cell line characteristics (16). In addition, whether infectivity is limited by the presence of virus-specific fecal antibodies and the possibility that cellular host factors may prevent or limit replication of certain genotypes all indicate that more research is needed to further optimize this cultivation system.
Dr. Costantini is an associate service fellow at the National Calicivirus Laboratory, Division of Viral Diseases, National Center for Immunization and Respiratory Diseases, Centers for Disease Control and Prevention in Atlanta, Georgia, USA. Her research interests include all aspects of viral gastroenteritis with an emphasis on norovirus immunology and vaccines.
Acknowledgment
This study was supported by the intramural food safety program at the Centers for Disease Control and Prevention and in part by Agriculture and Food Research Initiative Competitive Grant 2011-68003-30395 from the US Department of Agriculture, National Institute of Food and Agriculture and PHS grants P01AI 057788 and P30 DK 56338. Names of specific vendors, manufacturers, or products are included for public health and informational purposes; inclusion does not imply endorsement of the vendors, manufacturers, or products by the Centers for Disease Control and Prevention or the US Department of Health and Human Services.
References
- Ahmed SM, Hall AJ, Robinson AE, Verhoef L, Premkumar P, Parashar UD, et al. Global prevalence of norovirus in cases of gastroenteritis: a systematic review and meta-analysis. Lancet Infect Dis. 2014;14:725–30.DOIPubMedGoogle Scholar
- Duizer E, Schwab KJ, Neill FH, Atmar RL, Koopmans MP, Estes MK. Laboratory efforts to cultivate noroviruses. J Gen Virol. 2004;85:79–87.DOIPubMedGoogle Scholar
- Straub TM, Höner zu Bentrup K, Orosz-Coghlan P, Dohnalkova A, Mayer BK, Bartholomew RA, et al. In vitro cell culture infectivity assay for human noroviruses. Emerg Infect Dis. 2007;13:396–403.DOIPubMedGoogle Scholar
- Herbst-Kralovetz MM, Radtke AL, Lay MK, Hjelm BE, Bolick AN, Sarker SS, et al. Lack of norovirus replication and histo-blood group antigen expression in 3-dimensional intestinal epithelial cells. Emerg Infect Dis. 2013;19:431–8.DOIPubMedGoogle Scholar
- Papafragkou E, Hewitt J, Park GW, Greening G, Vinjé J. Challenges of culturing human norovirus in three-dimensional organoid intestinal cell culture models. PLoS One. 2013;8:e63485.DOIPubMedGoogle Scholar
- Takanashi S, Saif LJ, Hughes JH, Meulia T, Jung K, Scheuer KA, et al. Failure of propagation of human norovirus in intestinal epithelial cells with microvilli grown in three-dimensional cultures. Arch Virol. 2014;159:257–66.DOIPubMedGoogle Scholar
- Wobus CE, Karst SM, Thackray LB, Chang KO, Sosnovtsev SV, Belliot G, et al. Replication of Norovirus in cell culture reveals a tropism for dendritic cells and macrophages. PLoS Biol. 2004;2:e432.DOIPubMedGoogle Scholar
- Lay MK, Atmar RL, Guix S, Bharadwaj U, He H, Neill FH, et al. Norwalk virus does not replicate in human macrophages or dendritic cells derived from the peripheral blood of susceptible humans. Virology. 2010;406:1–11.DOIPubMedGoogle Scholar
- Jones MK, Watanabe M, Zhu S, Graves CL, Keyes LR, Grau KR, et al. Enteric bacteria promote human and mouse norovirus infection of B cells. Science. 2014;346:755–9.DOIPubMedGoogle Scholar
- Jones MK, Grau KR, Costantini V, Kolawole AO, de Graaf M, Freiden P, et al. Human norovirus culture in B cells. Nat Protoc. 2015;10:1939–47.DOIPubMedGoogle Scholar
- Karandikar UC, Crawford SE, Ajami NJ, Murakami K, Kou B, Ettayebi K, et al. Detection of human norovirus in intestinal biopsies from immunocompromised transplant patients. J Gen Virol. 2016;97:2291–300.DOIPubMedGoogle Scholar
- Sato T, Stange DE, Ferrante M, Vries RG, Van Es JH, Van den Brink S, et al. Long-term expansion of epithelial organoids from human colon, adenoma, adenocarcinoma, and Barrett’s epithelium. Gastroenterology. 2011;141:1762–72.DOIPubMedGoogle Scholar
- Saxena K, Blutt SE, Ettayebi K, Zeng XL, Broughman JR, Crawford SE, et al. Human intestinal enteroids: a new model to study human rotavirus infection, host restriction, and pathophysiology. J Virol. 2015;90:43–56.DOIPubMedGoogle Scholar
- VanDussen KL, Marinshaw JM, Shaikh N, Miyoshi H, Moon C, Tarr PI, et al. Development of an enhanced human gastrointestinal epithelial culture system to facilitate patient-based assays. Gut. 2015;64:911–20.DOIPubMedGoogle Scholar
- Zachos NC, Kovbasnjuk O, Foulke-Abel J, In J, Blutt SE, de Jonge HR, et al. Human enteroids/colonoids and intestinal organoids functionally recapitulate normal intestinal physiology and pathophysiology. J Biol Chem. 2016;291:3759–66.DOIPubMedGoogle Scholar
- Ettayebi K, Crawford SE, Murakami K, Broughman JR, Karandikar U, Tenge VR, et al. Replication of human noroviruses in stem cell-derived human enteroids. Science. 2016;353:1387–93.DOIPubMedGoogle Scholar
- Zou WY, Blutt SE, Crawford SE, Ettayebi K, Zeng XL, Saxena K, et al. Human intestinal enteroids: new models to study gastrointestinal virus infections. Methods Mol Biol. 2017.DOIPubMedGoogle Scholar
- Leon JS, Kingsley DH, Montes JS, Richards GP, Lyon GM, Abdulhafid GM, et al. Randomized, double-blinded clinical trial for human norovirus inactivation in oysters by high hydrostatic pressure processing. Appl Environ Microbiol. 2011;77:5476–82.DOIPubMedGoogle Scholar
- Keswick BH, Satterwhite TK, Johnson PC, DuPont HL, Secor SL, Bitsura JA, et al. Inactivation of Norwalk virus in drinking water by chlorine. Appl Environ Microbiol. 1985;50:261–4.PubMedGoogle Scholar
- Hoelzer K, Fanaselle W, Pouillot R, Van Doren JM, Dennis S. Virus inactivation on hard surfaces or in suspension by chemical disinfectants: systematic review and meta-analysis of norovirus surrogates. J Food Prot. 2013;76:1006–16.DOIPubMedGoogle Scholar
- Zonta W, Mauroy A, Farnir F, Thiry E. Comparative virucidal efficacy of seven disinfectants against murine norovirus and feline calicivirus, surrogates of human norovirus. Food Environ Virol. 2016;8:1–12.PubMedGoogle Scholar
- Mormann S, Heißenberg C, Pfannebecker J, Becker B. Tenacity of human norovirus and the surrogates feline calicivirus and murine norovirus during long-term storage on common nonporous food contact surfaces. J Food Prot. 2015;78:224–9.DOIPubMedGoogle Scholar
- Bozkurt H, D’Souza DH, Davidson PM. Thermal inactivation of foodborne enteric viruses and their viral surrogates in foods. J Food Prot. 2015;78:1597–617.DOIPubMedGoogle Scholar
- Cromeans T, Park GW, Costantini V, Lee D, Wang Q, Farkas T, et al. Comprehensive comparison of cultivable norovirus surrogates in response to different inactivation and disinfection treatments. Appl Environ Microbiol. 2014;80:5743–51.DOIPubMedGoogle Scholar
- Cannon JL, Papafragkou E, Park GW, Osborne J, Jaykus LA, Vinjé J. Surrogates for the study of norovirus stability and inactivation in the environment: aA comparison of murine norovirus and feline calicivirus. J Food Prot. 2006;69:2761–5.DOIPubMedGoogle Scholar
- Cannon JL, Barclay L, Collins NR, Wikswo ME, Castro CJ, Magaña LC, et al. Genetic and epidemiologic trends of norovirus outbreaks in the United States from 2013 to 2016 demonstrated emergence of novel GII.4 recombinant viruses. J Clin Microbiol. 2017;55:2208–21.DOIPubMedGoogle Scholar
- Vega E, Barclay L, Gregoricus N, Shirley SH, Lee D, Vinjé J. Genotypic and epidemiologic trends of norovirus outbreaks in the United States, 2009 to 2013. J Clin Microbiol. 2014;52:147–55.DOIPubMedGoogle Scholar
- Reed LJ, Muench H. A simple method of estimating fifty per cent endpoints. Am J Hyg. 1938;27:493–97.
- Drummond CG, Bolock AM, Ma C, Luke CJ, Good M, Coyne CB. Enteroviruses infect human enteroids and induce antiviral signaling in a cell lineage-specific manner. Proc Natl Acad Sci U S A. 2017;114:1672–7.DOIPubMedGoogle Scholar
- Saxena K, Simon LM, Zeng XL, Blutt SE, Crawford SE, Sastri NP, et al. A paradox of transcriptional and functional innate interferon responses of human intestinal enteroids to enteric virus infection. Proc Natl Acad Sci U S A. 2017;114:E570–9.DOIPubMedGoogle Scholar
- Kingsley DH, Fay JP, Calci K, Pouillot R, Woods J, Chen H, et al. Evaluation of chlorine treatment levels for inactivation of human norovirus and MS2 bacteriophage during sewage treatment. Appl Environ Microbiol. 2017;83:e01270-17.DOIPubMedGoogle Scholar
- Yin X, Farin HF, van Es JH, Clevers H, Langer R, Karp JM. Niche-independent high-purity cultures of Lgr5+ intestinal stem cells and their progeny. Nat Methods. 2014;11:106–12.DOIPubMedGoogle Scholar
Figures
Tables
Cite This ArticleTable of Contents – Volume 24, Number 8—August 2018
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Veronica Costantini, Centers for Disease Control and Prevention, 1600 Clifton Rd NE, Mailstop G04, Atlanta, GA 30329-4027, USA
Top