Volume 31, Number 3—March 2025
Research
Influenza A(H5N1) Immune Response among Ferrets with Influenza A(H1N1)pdm09 Immunity
Cite This Article
Citation for Media
Abstract
The emergence of highly pathogenic avian influenza A(H5N1) virus in dairy cattle herds across the United States in 2024 caused several human infections. Understanding the risk for spillover infections into humans is crucial for protecting public health. We investigated whether immunity from influenza A(H1N1)pdm09 (pH1N1) virus would provide protection from death and severe clinical disease among ferrets intranasally infected with H5N1 virus from dairy cows from the 2024 outbreak. We observed differential tissue tropism among pH1N1-immune ferrets. pH1N1-immune ferrets also had little H5N1 viral dissemination to organs outside the respiratory tract and much less H5N1 virus in nasal secretions and the respiratory tract than naive ferrets. In addition, ferrets with pH1N1 immunity produced antibodies that cross-reacted with H5N1 neuraminidase protein. Taken together, our results suggest that humans with immunity to human seasonal influenza viruses may experience milder disease from the 2024 influenza A(H5N1) virus strain.
In March 2024, an outbreak of highly pathogenic avian influenza A(H5N1) clade 2.3.4.4b virus was identified in dairy cattle herds in Texas, USA, and then spread to >400 herds in >15 states (1). That spread emphasized the need to monitor H5N1 clade 2.3.4.4b for pandemic potential. H5N1 clade 2.3.4.4b virus infections of various mammals resulted in severe disease and death, including among foxes, mink, cats, cetaceans, pinnipeds, and cows (2,3). In early April 2024, a case of human infection was identified in Texas (4), and more human H5N1 cases were identified among workers associated with poultry or dairy farms in California, Missouri, Michigan, Colorado, and Washington (5). In August 2024, human infections in the United States were characterized by conjunctivitis and mild respiratory symptoms, and most did not require hospitalization (5).
Most persons experience their first influenza virus infection by 5 years of age (6). Thus, current H5N1 human infections are occurring among persons with prior influenza A virus (IAV) immunity. The reduced disease severity among persons infected with the 2024 H5N1 virus might be driven by immunity to human seasonal influenza viruses. Statistical modeling analysis of known human cases of H5N1 and H7N9 infection indicated that childhood hemagglutinin (HA) imprinting may provide lifelong protection against severe infection and death from those viruses (7). Specifically, previous research has suggested that immune imprinting with human seasonal H1N1 or H2N2 influenza viruses would reduce disease severity to H5N1 because H5, H1, and H2 share a similar group 1 HA stalk domain (7). Despite the potential effects such immunity could have to reduce H5N1 replication and pathogenesis, risk assessment of the 2024 H5N1 outbreak strain has only been performed in immunologically naive ferrets (8). We investigated whether ferrets with H1N1 immunity would experience reduced virus replication and disease severity from dairy cow H5N1 virus.
Cell Preparation
We obtained MDCK cells and human 293T cells from the American Type Culture Collection (https://www.atcc.org). We maintained the MDCK cells in minimum essential medium and the 293T cells in Dulbecco Modified Eagle Medium. We supplemented both cell media with 10% fetal bovine serum, 2 mmol L-glutamine, 100 U/mL penicillin, and 100 mg/mL streptomycin. We incubated cells at 37°C with 5% carbon dioxide (CO2). We obtained human 293F cells Medium (Thermo Fisher Scientific, https://www.fishersci.com) and maintained cells at 37°C with 5%–8% CO2 in FreeStyle 293 Expression (Thermo Fisher Scientific) supplemented with 100 U/mL penicillin and 100 mg/mL streptomycin.
Virus Generation
We generated A/dairy cattle/Texas/24-008749-001/2024(H5N1) (GISAID accession no. EPI_ISL_19014384) virus from 8 plasmid reverse genetics system and propagated in MDCK cells (Appendix). We determined noncoding regions for each segment from consensus alignment of H5N1 strains from the 2.3.4.4b clade viruses.
Human Subjects Research and Ethics Statement
Figure 1
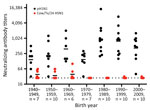
Figure 1. Neutralizing antibody titers in human serum used in a study of influenza A(H5N1) immune response among ferrets with pH1N1 immunity. We used serum samples collected from healthy persons during 2020–2021...
As part of this research, we assessed human serum samples for cross-reactive antibodies to H5N1 virus (Figure 1). The University of Pittsburgh institutional review board approved collection of serum samples from healthy adult donors who provided written informed consent for their samples to be used in infectious disease research (protocol approval no. STUDY20030228). All participants self-reported age, sex, ethnicity, and race.
Ferret Infections
We screened ferrets before this study to ensure no influenza immunity before they arrived at University of Pittsburgh (Appendix). Using our previously developed preimmune ferret model (9,10), we infected 5 ferrets with recombinant influenza A(H1N1)pdm09 (pH1N1) virus by using the A/California/07/2009 strain. We infected ferrets either experimentally by intranasal introduction of A/California/07/2009 or naturally by exposure to an experimentally infected ferret in a controlled transmission study conducted at the University of Pittsburgh.
pH1N1-immune animals then recovered from acquired infections and were housed for 98 days before we infected 5 pH1N1-immune and 5 immunologically naive ferrets with A/dairy cattle/Texas/24-008749-001/2024(H5N1), termed cow/Tx/24 H5N1. We intranasally inoculated all 10 ferrets with 104 50% tissue culture infectious dose (TCID50) cow/Tx/24 H5N1 virus in 500 µL of L-15 media (250 µL in each nostril). We monitored ferrets daily during the postinoculation period and recorded clinical signs, including weight loss, temperature, activity, sneezing, coughing, and nasal discharge, as previously described (11). For animals that reached >10% weight loss, we provided urgent care diet cat food 2×/day to entice eating.
Figure 2
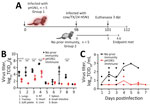
Figure 2. Infection timeline and virus replication titers in a study of influenza A(H5N1) immune response among ferrets with pH1N1 immunity. A) Schematic of experimental timeline for 2 groups of ferrets intranasally...
Humane endpoints for this study included bodyweight loss >20% (relative to weight at challenge) and a prolonged inactivity as assessed by trained veterinary staff. Three animals from each group were euthanized 3 days postinoculation (dpi) for tissue titration; the other 2 ferrets from each group were kept for >14 days or until they reached endpoint criteria (Figure 2, panel A).
Ferret Sample Collection and Preparation
We collected nasal wash from each ferret at 1–7 days postinoculation (dpi). To examine whether pH1N1 altered cow/Tx/24 H5N1 tissue tropism, we euthanized 3 intranasally infected ferrets from each group at 3 dpi to collect tissues (lungs, trachea, soft palate, nasal turbinates, heart, liver, spleen, small intestine, and brain) and determined virus titers (Appendix).
We titered nasal wash and organ samples in MDCK cell cultures. We made 10-fold serial dilutions and inoculated dilutions on 96-well plates by using 4 wells/dilution. We observed the MDCK cells at 4 dpi for cytopathic effect (CPE). We calculated viral titers by using the Reed and Muench method (12) and expressed the results as log10 TCID50/mL.
Animal Ethics Statement
Ferret experiments were conducted in Biosecurity Level 2 and 3 facilities at the University of Pittsburgh in compliance with the guidelines of the Institutional Animal Care and Use Committee (approved protocol nos. 22061230 and 21089461). For all nasal washes and survival blood draws, animals were sedated with isoflurane following approved methods. Ketamine and xylazine were used for sedation for all terminal procedures, followed by cardiac administration of euthanasia solution. Approved University of Pittsburgh Division of Laboratory Animal Resources staff administered euthanasia. H5N1 studies were performed in accordance with the University of Pittsburgh select agent permit no. 20230320-074008.
Microneutralization Assays
We heat inactivated human and ferret serum samples at 56°C for 30 minutes. We determined the titer of neutralizing antibodies by incubating 2-fold serial dilutions of the heat-inactivated serum samples with 103.3 TCID50 of influenza virus for 1 hour at room temperature with continuous rocking. We added media with tosyl phenylalanyl chloromethyl ketone–treated trypsin to 96-well plates with confluent MDCKs before adding the virus-serum mixture. After 4 days, we determined the CPE and expressed the neutralizing antibody titer as the reciprocal of the highest dilution of serum required to completely neutralize the infectivity of each virus on MDCK cells. We calculated the concentration of antibody required to neutralize 100 TCID50 of virus on the basis of the neutralizing titer dilution divided by the initial dilution factor, multiplied by the antibody concentration.
Histology
We stained respiratory tissue sections collected from euthanized ferrets with hematoxylin and eosin for histopathologic analysis or influenza A nucleoprotein for immunohistochemistry (Appendix). We initially examined the prepared slides blinded to experimental groups to eliminate observer bias, then by unblinding for figure preparation. We developed an ordinal scoring system to summarize the histopathologic and immunohistochemical findings: 0, not observed; 1 (mild), <10% of parenchyma impacted; 2 (moderate), from 10%–25% of parenchyma affected; and 3 (severe), 25%–50% of parenchyma affected.
ELISA
We adhered 500 ng of recombinant HA full-length soluble ectodomains or recombinant neuraminidase (NA) to high-capacity binding 96 well-plates (Corning, https://www.corning.com) overnight in phosphate-buffered saline (PBS) at 4°C (Appendix). We then washed the HA- or NA-coated plates with a 0.05% vol/vol PBS-Tween-20 (PBS-T) buffer and then blocked with PBS-T containing 2% bovine serum albumin for 1 hour at room temperature. We removed the blocking solution and added 2-fold dilutions of ferret serum in blocking solution to the wells. We then incubated the plates for 1 hour at room temperature. We removed the primary antibody solution and washed the plates 3 times with PBS-T. We added a secondary Goat Anti-Ferret IgG H&L (HRP) (Abcam, https://www.abcam.com) diluted 1:10,000 in blocking solution to the wells and incubated for 30 minutes at room temperature. We then washed the plates 3 times with PBS-T. We developed the plates by using 150 μL 1-Step TMB Substrate (Thermo Fisher Scientific). After a brief incubation at room temperature, we stopped HRP reactions by adding 100 μL of 4N sulfuric acid solution. We read the plates by using a SpectraMax 340PC384 Microplate Reader (Molecular Devices, https://www.moleculardevices.com) at 450 nm. We performed all measurements in duplicate. We then graphed the average of the 2 measurements for each ferret sample as the mean absorbance at 450 nm by using Prism software version 9.0 (GraphPad, https://www.graphpad.com).
Neutralizing Antibody Levels in Humans
H5N1 IAVs have not circulated widely in the human population, and major immunity against those strains likely does not exist. To assess whether any cross-reactive antibodies existed in the human population, we conducted neutralization assays with human serum against cow/Tx/24 H5N1 and pH1N1, and results revealed high levels of circulating antibodies against pH1N1 in persons of all ages (Figure 1). Of note, 12 of the 60 serum samples tested had detectable levels of cross-neutralizing antibodies against cow/Tx/24 H5N1 that were above the limit of detection. Of the 12 serum samples with cross-neutralizing antibodies, 10 were collected from persons born in the 1940s, 1950s, and 1960s and 2 were from persons born after 1970 (Figure 1), which correlates well with H5 cross-reactive antibodies in older persons (13). Those data suggests persons born after 1980 could be more susceptible to infection with H5N1 virus from dairy cows. We do not know the ages of persons with documented H5N1 infections since 2022.
Effect of pH1N1 Immunity on Viral Titers and Dissemination
We sought to extend our prior work (9,10) and examine the role of pH1N1 immunity on dairy cow H5N1 infection severity and replication in the ferret model. In ferrets without pH1N1 immunity, cow/Tx/24 H5N1 resulted in high viral loads in the respiratory tissues and produced a systemic infection, as observed by virus detection in the heart, liver, spleen, and intestine (Figure 2, panel B). In contrast, ferrets with pH1N1 immunity exhibited lower levels of virus replication that were limited to the respiratory tract and were statistically significant (p<0.01) (Figure 2, panel B). The lack of virus in the brain of ferrets without pH1N1 immunity at 3 dpi is consistent with data reported from other groups (8).
Nasal wash titers were also drastically different between the 2 groups of ferrets. Virus was consistently detected over time in the nasal washes of ferrets without prior immunity, whereas most pH1N1-immune ferrets had no detectable cow/Tx/24 H5N1 virus in nasal washes; the exceptions were 1 ferret at 4 dpi and a different ferret at 6 dpi (Figure 2, panel C). Of note, we detected virus in the nasal turbinates of the pH1N1-immune ferrets euthanized at 3 dpi, despite a lack of virus in the nasal wash; however, virus levels were much lower than among animals without prior immunity (Figure 2, panels B, C). That difference could be attributed to the methods of sample collection. Nasal washes are performed by pushing fluid through 1 nostril and collecting the liquid from the other nostril, which samples the tip of the turbinates. In contrast, the entire nasal turbinate tissue is collected at the time of necropsy and homogenized to collect any released and cell associated viruses; thus, turbinates would be expected to have higher virus levels.
Figure 3
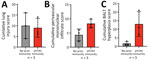
Figure 3. Lung infiltrates measured in a study of influenza A(H5N1) immune response among ferrets with pH1N1 immunity. We blindly scored 5 lung sections for ferrets with no prior or existing pH1N1...
Figure 4
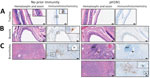
Figure 4. Hematoxylin and eosin–stained and immunohistochemistry tissue samples from a study of influenza A(H5N1) immune response among ferrets with pH1N1 immunity. A) Tracheal tissue. Scale bars indicate 20 mm; inset shows...
Histopathological analysis of lung tissues harvested at 3 dpi indicated both groups of ferrets had similar lung injury (Figure 3, panel A). However, more detailed examination of the data indicated that ferrets with pH1N1 immunity had more residual mononuclear perivascular infiltrates and bronchus-associated lymphoid tissue (BALT) hyperplasia (Figure 3, panels B, C), which may play a role in preventing development of severe clinical disease. Immunohistochemistry with IAV nucleoprotein (NP) indicated that pH1N1-immune ferrets had limited NP-positive cells in the trachea, mainstem bronchi, and bronchioles than ferrets without prior immunity (Figure 4). In pH1N1-immune ferrets with areas of BALT hyperplasia, we detected limited viral antigen and necrotizing bronchointerstitial pneumonia (Figure 4, panel C). Furthermore, we observed IAV NP antigen in alveolar pneumocytes (both type 1 and 2) in ferrets irrespective of immune status (Figure 4, panel C). Those findings are in contrast to pH1N1 infection in ferrets, in which pH1N1 virus infected epithelial cells in the large and small airways (14–18).
Figure 5
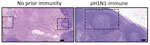
Figure 5. Hematoxylin and eosin–stained lymph node samples from a study of influenza A(H5N1) immune response among ferrets with pH1N1 immunity. Ferrets with no prior immunity (left panel) or pH1N1 (right panel)...
Examination of the tracheobronchial lymph node histology revealed more lymphoid depletion, necrosis, fibrin, and edema in ferrets without pH1N1 immunity compared with pH1N1-immune ferrets (Figure 5). Overall, those data indicated that resident lymphoid changes in ferrets with pH1N1 immunity may have reduced cow/Tx/24 H5N1 replication and dissemination to other organs, which could affect disease severity.
Effects of pH1N1 Immunity on H5N1 Mortality and Severe Disease
Figure 6

Figure 6. Mortality rates and weight change in a study of influenza A(H5N1) immune response among ferrets with pH1N1 immunity. A) Mortality rates; B) percentage weight change. We observed percentage of weight...
Figure 7
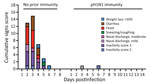
Figure 7. Cumulative clinical signs scores in a study of influenza A(H5N1) immune response among ferrets with pH1N1 immunity. Ferrets with or without pH1N1 immunity were intranasally infected with H5N1 strain A/dairy...
We followed H5N1-infected ferrets with (n = 2) and without (n = 2) pH1N1 immunity to 14 dpi to examine death outcomes (Figure 6, panel A). The 2 animals with pH1N1 immunity survived challenge with cow/Tx/24 H5N1 virus, whereas the 2 immunologically naive ferrets were humanly euthanized at 4 dpi and 6 dpi because severe clinical signs developed (Figure 6, panel A). We observed that ferrets with pH1N1 immunity had <5% weight loss, whereas naive ferrets experienced >10% weight loss (Figure 6, panel B). Assessment of clinical signs, such as diarrhea, fever, nasal discharge and playfulness, revealed more severe clinical signs in all immunologically naive animals than among those with prior pH1N1 immunity (Figure 7).
Surviving ferrets with pH1N1 immunity seroconverted against cow/Tx/24 H5N1, albeit to low microneutralization titers of 20 and 80 (Table). In addition, neither of the 2 pH1N1-immune ferrets had a >4-fold rise in pH1N1 antibodies after cow/Tx/24 H5N1 challenge (Table). Taken together, those data indicated that pH1N1 immunity protects ferrets from severe clinical disease and death caused by cow/Tx/24 H5N1 infection.
Cross-Reactive NA Antibody Production
Figure 8

Figure 8. Cross-reactive NA binding antibodies in a study of influenza A(H5N1) immune response among ferrets with pH1N1. Ferrets with or without pH1N1 immunity were intranasally infected with H5N1 strain A/dairy cattle/Texas/24-008749-001/2024(H5N1)....
IAV infection induces antibody responses against HA and NA proteins that can provide varying levels of protection against subsequent infections (19). In addition, cross-reactive HA stalk-specific antibodies are able to play a role in reducing influenza virus disease severity (20–22). To identify immune factors that contribute to the protection of pH1N1-immune ferrets from severe disease, we measured neutralizing and total HA binding antibodies. Before challenge with cow/Tx/24 H5N1 virus, ferrets with pH1N1 immunity exhibited high levels of neutralizing antibodies against pH1N1 but no neutralizing antibodies above the limit of detection against cow/Tx/24 H5N1 (Figure 8, panel A).
To explore the production of nonneutralizing cross-reactive HA antibodies, we performed an ELISA with serum from ferrets with pH1N1 immunity by using the whole H1 (A/California/07/2009 H1N1) or H5 (A/dairy cattle/Texas/24008749001/2024 H5N1) HA protein (Figure 8, panel B). Ferrets with pH1N1 immunity produced antibodies that bound to H1 as expected but displayed the same background levels of antibody binding to the H5 HA protein as ferrets with no prior immunity (Figure 8, panel B), indicating no detectable cross-reactive HA antibodies against the avian H5 protein.
Finally, we performed an ELISA using NA from a human (A/Michigan/45/2015 H1N1) or avian (A/mallard/New York/22-008760–007-original/2022 H5N1, which is 98.7% similar to cow/Tx/24 NA) IAV to determine whether pH1N1-immune ferrets had any cross-reacting NA antibodies that might contribute to the protection against severe disease before challenge with H5N1. Of note, serum samples from pH1N1-immune ferrets had antibodies that bound to both the human and avian NA antigens, but ferrets without immunity had background binding levels (Figure 8, panel C). Those data suggest that cross-reactive NA antibodies to avian N1 may be produced from a human seasonal pH1N1 infection.
Influenza A(H1N1)pdm09 immunity in ferrets was sufficient to protect from severe disease and death from highly pathogenic avian influenza A(H5N1) virus from dairy cows. We also observed significantly reduced H5N1 viral titers in nasal secretions and respiratory tract tissues in the animals with pH1N1 immunity (p<0.01). Of note, protection from H5N1 infection was not due to cross-neutralizing antibodies in serum because ferrets with pH1N1 immunity did not generate systemic antibodies that cross-neutralized the cow/Tx/24 H5N1 virus (Figure 8, panel A). Rather, we found ferrets with pH1N1 immunity produced cross-reacting antibodies to H5N1 NA protein (Figure 8, panel C), which is consistent with observations reported from human serologic data (23). Immunity to NA has previously been implicated in providing protection during the 1968 H3N2 pandemic (24,25), and can reduce disease severity of naturally infected and experimentally challenged persons (26).
NA antibodies may be involved in protection from severe disease observed in the pH1N1-immune ferrets. However, further studies on the mechanisms of protection are clearly warranted and should include an examination of mucosal immunity from antibodies in the respiratory tract that have broad binding potential. Tissue-resident memory T cells may also help reduce the severity of disease, as is suspected in the case of H1 immunity protecting from airborne transmission of human seasonal H3N2 virus (9). A conservation of immunodominant T-cell epitopes between H5N1 and seasonal influenza viruses, including H1N1, was recently reported and suggested to potentially provide a level of cross-protective immunity (27). We did note that the lung tissues of ferrets with pH1N1 immunity had increased mononuclear perivascular infiltrates and BALT hyperplasia, consistent with tissue-specific T-cell responses, although additional investigation is required.
The mild infection noted in the 2 pH1N1-immune ferrets that survived until day 14 might account for the low levels of neutralizing antibodies against cow/Tx/24 H5N1 (Table). That observation may be critical to inform the use of H5 seroconversion as a detection mechanism for prevalence of H5 infections in farm workers because mild infections may not produce a robust systemic antibody immune response.
All adults have immunity from repeated influenza virus infections over their lifetimes, but how previous exposures translate into protection may be strain-dependent and change over time. Human H5N1 infections during 2003 had a 30%–50% mortality rate worldwide (28). However, since the emergence of the 2.3.4.4b clade in 2020, the mortality rate has been declining; during 2020–2024, at least 80 human infections with various H5N1 clades were reported, but only 8 deaths were reported (28). Of note, in 2024, at least 72 human infections and 2 deaths from H5N1 have occurred; both deaths were reported in Cambodia from 2.3.2.1c clade, which is distinct from 2.3.4.4b (28). The mild clinical manifestations of H5N1 human cases in the United States could be due to several factors, including changes in the viral genome that result in a less pathogenic virus, inoculation routes and doses, immunity to H1N1 strains that circulated widely since 2010, or a combination of those factors. However, additional research into the level of protection afforded by other human seasonal influenza viruses, particularly currently circulating H1N1 viruses and those before the 2009 H1N1 pandemic, is needed to assess whether currently circulating H1N1 viruses produce a protective immune signature but other prior strains do not. In particular, understanding whether infection from H1N1 or H2N2 strains circulating before 1970 can produce antibodies that cross-react with H5N1 would be useful, given the presence of cross-neutralizing antibodies observed persons born before 1970. Finally, determining whether immunity to conserved regions of the NA or other viral proteins are driving the protection observed with pH1N1 infection is crucial because we detected no neutralizing antibodies in younger persons (Figure 1).
One limitation of this study is the small number of human serum samples used to test for cross-neutralizing antibodies against dairy cow H5N1. However, another group reported similar findings from a different assay (T.A. Garretson et al., unpub. data, https://doi.org/10.1101/2024.10.31.24316514). Other limitations that should be addressed in future work include the small number of animals used in our preimmune studies, use of only 1 subtype of human seasonal virus for the immune imprint, and challenge with only 1 strain of H5N1. Lethality after highly pathogenic avian A(H5N1) influenza infection can be strain specific, and published work has shown that another dairy cattle H5N1 strain was only partially lethal in immunologically naive ferrets (30), but a human H5N1 isolate from Texas was lethal in ferrets (29,30). Therefore, assessment of additional H5N1 strains isolated from dairy cows and human spillover infections is needed. Other groups have performed studies with ferrets having imprints from vaccination or seasonal human influenza virus infection and shown protection against different H5N1 strains (31–35; P.H. Brigleb et al., unpub. data, https://doi.org/10.1101/2024.10.23.619695).
In conclusion, we found ferrets with immunity to pH1N1 virus exhibited reduced H5N1 virus replication and dissemination, had less mortality and fewer disease symptoms from H5N1 infection, and expressed H5N1 cross-reacting antibodies to the NA protein. Those results suggest that immunity to heterotypic influenza viruses may explain the mild symptoms observed during 2024 H5N1 infection of dairy and poultry farm workers. Although human H5N1 infections from the 2024 outbreak resulted in mostly mild illnesses, additional research addressing the effects of prior influenza immunity on the pathogenesis and transmission of H5N1 could shed light on the 2024 outbreak strain and inform pandemic risk plans.
This article was preprinted at https://www.biorxiv.org/content/10.1101/2024.10.23.619881v1.
Dr. Le Sage is a research assistant professor at the University of Pittsburgh Center for Vaccine Research. Her research interests include elucidating the requirements for influenza virus transmission and assessing the pandemic potential of emerging influenza viruses.
Acknowledgments
We would like to extend our deepest gratitude to Julian Arthur and Jessica Simendinger for their outstanding work in developing the Influenza A Nucleoprotein (NP) (F8L6X) Rabbit mAb #99797 (Cell Signaling Technology, Inc., https://www.cellsignal.com). Their expertise, dedication, and tireless efforts have been instrumental in the successful creation of this critical clone, and we are particularly grateful for their collaborative efforts with our team during the validation process. We thank Rachel Duron for critical review and feedback.
All source data are available on Figshare (https://doi.org/10.6084/m9.figshare.25843414).
This project has been funded in part with federal funds from the National Institute of Allergy and Infectious Diseases (NIAID), National Institutes of Health (NIH), Department of Health and Human Services, under contract no. 75N93021C00015. A.K.M. receives funding from Burroughs Wellcome Career Awards for Medical Scientists (award no. 1013362.02). The University of Pittsburgh Regional Biocontainment Laboratory within the Center for Vaccine Research is supported by an NIH award (no. UC7AI180311) from NIAID. N.A.C. is supported by NIH S10 instrumentation awards (nos. S10OD030269 and S10OD026983). Work in the Krammer laboratory was supported via an NIAID-funded Center of Excellence for Influenza Research and Response contract (no. 75N93021C00014).
The Icahn School of Medicine at Mount Sinai has filed patent applications relating to influenza virus vaccines and therapeutic vaccines which list F.K. as coinventor. Several of those patents have been licensed and F.K. has received royalty payments from commercial entities. F.K. has consulted for Merck, Pfizer, Seqirus, GlaxoSmithKline, and Curevac and is currently consulting for Gritstone, 3rd Rock Ventures, and Avimex, and he is a cofounder and scientific advisory board member of CastleVax. The Krammer laboratory is also collaborating with Dynavax on influenza virus vaccine development and with VIR on influenza therapeutics. All other authors declare no competing financial or nonfinancial interests in relation to the work described.
Author contributions: V.L. and S.S.L. designed the experiments, analyzed, interpreted the data, and wrote the manuscript. V.L., B.D.W., G.A.M., S.E.P., A.K.O., H.C.S., and N.A.C. performed the experiments. K.R.M., D.S.R., L.H.M., D.B., F.K., A.K.M., and W.P.D. contributed resources and analysis. All authors edited and approved the manuscript.
References
- US Department of Agriculture. HPAI confirmed cases in livestock [cited 2024 Nov 9]. https://www.aphis.usda.gov/livestock-poultry-disease/avian/avian-influenza/hpai-detections/hpai-confirmed-cases-livestock
- Koopmans MPG, Barton Behravesh C, Cunningham AA, Adisasmito WB, Almuhairi S, Bilivogui P, et al.; One Health High-Level Expert Panel. The panzootic spread of highly pathogenic avian influenza H5N1 sublineage 2.3.4.4b: a critical appraisal of One Health preparedness and prevention. Lancet Infect Dis. 2024;24:e774–81.
- Peacock TP, Moncla L, Dudas G, VanInsberghe D, Sukhova K, Lloyd-Smith JO, et al. The global H5N1 influenza panzootic in mammals. Nature. 2025;637:304–13.
- Uyeki TM, Milton S, Abdul Hamid C, Reinoso Webb C, Presley SM, Shetty V, et al. Highly pathogenic avian influenza A(H5N1) virus infection in a dairy farm worker. N Engl J Med. 2024;390:2028–9.
- Centers for Disease Control and Prevention. Avian influenza (bird flu) [cited 2024 Nov 9]. https://www.cdc.gov/bird-flu/spotlights/ah5n1-response-update.html
- Bodewes R, de Mutsert G, van der Klis FR, Ventresca M, Wilks S, Smith DJ, et al. Prevalence of antibodies against seasonal influenza A and B viruses in children in Netherlands. Clin Vaccine Immunol. 2011;18:469–76.
- Gostic KM, Ambrose M, Worobey M, Lloyd-Smith JO. Potent protection against H5N1 and H7N9 influenza via childhood hemagglutinin imprinting. Science. 2016;354:722–6.
- Eisfeld AJ, Biswas A, Guan L, Gu C, Maemura T, Trifkovic S, et al. Pathogenicity and transmissibility of bovine H5N1 influenza virus. Nature. 2024;633:426–32.
- Le Sage V, Jones JE, Kormuth KA, Fitzsimmons WJ, Nturibi E, Padovani GH, et al. Pre-existing heterosubtypic immunity provides a barrier to airborne transmission of influenza viruses. PLoS Pathog. 2021;17:
e1009273 . - Le Sage V, Rockey NC, French AJ, McBride R, McCarthy KR, Rigatti LH, et al. Potential pandemic risk of circulating swine H1N2 influenza viruses. Nat Commun. 2024;15:5025.
- Mueller Brown K, Le Sage V, French AJ, Jones JE, Padovani GH, Avery AJ, et al. Secondary infection with Streptococcus pneumoniae decreases influenza virus replication and is linked to severe disease. FEMS Microbes. 2022;3:
xtac007 . - Reed LJ, Muench H. A simple method of estimating fifty per cent endpoints. Am J Epidemiol. 1938;27:493–7.
- Nachbagauer R, Choi A, Izikson R, Cox MM, Palese P, Krammer F. Age dependence and isotype specificity of influenza virus hemagglutinin stalk-reactive antibodies in humans. MBio. 2016;7:e01996–15.
- Camp JV, Bagci U, Chu YK, Squier B, Fraig M, Uriarte SM, et al. Lower respiratory tract infection of the ferret by 2009 H1N1 pandemic influenza A virus triggers biphasic, dystemic, and local recruitment of neutrophils. J Virol. 2015;89:8733–48.
- Lakdawala SS, Jayaraman A, Halpin RA, Lamirande EW, Shih AR, Stockwell TB, et al. The soft palate is an important site of adaptation for transmissible influenza viruses. Nature. 2015;526:122–5.
- Lakdawala SS, Shih AR, Jayaraman A, Lamirande EW, Moore I, Paskel M, et al. Receptor specificity does not affect replication or virulence of the 2009 pandemic H1N1 influenza virus in mice and ferrets. Virology. 2013;446:349–56.
- Smith JH, Nagy T, Driskell E, Brooks P, Tompkins SM, Tripp RA. Comparative pathology in ferrets infected with H1N1 influenza A viruses isolated from different hosts. J Virol. 2011;85:7572–81.
- Vidaña B, Martínez J, Martínez-Orellana P, García Migura L, Montoya M, Martorell J, et al. Heterogeneous pathological outcomes after experimental pH1N1 influenza infection in ferrets correlate with viral replication and host immune responses in the lung. Vet Res (Faisalabad). 2014;45:85.
- Kosik I, Yewdell JW. Influenza hemagglutinin and neuraminidase: yin-yang proteins coevolving to thwart immunity. Viruses. 2019;11:346.
- Pica N, Hai R, Krammer F, Wang TT, Maamary J, Eggink D, et al. Hemagglutinin stalk antibodies elicited by the 2009 pandemic influenza virus as a mechanism for the extinction of seasonal H1N1 viruses. Proc Natl Acad Sci U S A. 2012;109:2573–8.
- Throsby M, van den Brink E, Jongeneelen M, Poon LL, Alard P, Cornelissen L, et al. Heterosubtypic neutralizing monoclonal antibodies cross-protective against H5N1 and H1N1 recovered from human IgM+ memory B cells. PLoS One. 2008;3:
e3942 . - Wu NC, Wilson IA. Influenza hemagglutinin structures and antibody recognition. Cold Spring Harb Perspect Med. 2020;10:
a038778 . - Daulagala P, Cheng SMS, Chin A, Luk LLH, Leung K, Wu JT, et al. Avian influenza A(H5N1) neuraminidase inhibition antibodies in healthy adults after exposure to influenza A(H1N1)pdm09. Emerg Infect Dis. 2024;30:168–71.
- Monto AS, Kendal AP. Effect of neuraminidase antibody on Hong Kong influenza. Lancet. 1973;301:623–5.
- Murphy BR, Kasel JA, Chanock RM. Association of serum anti-neuraminidase antibody with resistance to influenza in man. N Engl J Med. 1972;286:1329–32.
- Maier HE, Nachbagauer R, Kuan G, Ng S, Lopez R, Sanchez N, et al. Pre-existing antineuraminidase antibodies are associated with shortened duration of influenza A(H1N1)pdm virus shedding and illness in naturally infected adults. Clin Infect Dis. 2020;70:2290–7.
- Sidney J, Kim A-R, de Vries RD, Peters B, Meade PS, Krammer F, et al. Targets of influenza human T cell response are mostly conserved in H5N1. [Epub ahead of print]. MBio. 2024.
- World Health Organization. Cumulative number of confirmed human cases for avian influenza A(H5N1) reported to WHO, 2003–2024, 26 February 2024. [cited 2025 Feb 3] https://www.who.int/publications/m/item/cumulative-number-of-confirmed-human-cases-for-avian-influenza-a(h5n1)-reported-to-who--2003-2024--20-december-2024
- Eisfeld AJ, Biswas A, Guan L, Gu C, Maemura T, Trifkovic S, et al. Pathogenicity and transmissibility of bovine H5N1 influenza virus. Nature. 2024;633:426–32.
- Gu C, Maemura T, Guan L, Eisfeld AJ, Biswas A, Kiso M, et al. A human isolate of bovine H5N1 is transmissible and lethal in animal models. Nature. 2024;636:711–8.
- Pulit-Penaloza JA, Belser JA, Brock N, Kieran TJ, Sun X, Pappas C, et al. Transmission of a human isolate of clade 2.3.4.4b A(H5N1) virus in ferrets. Nature. 2024;636:705–10.
- Nuñez IA, Jang H, Huang Y, Kelvin A, Ross TM. Influenza virus immune imprinting dictates the clinical outcomes in ferrets challenged with highly pathogenic avian influenza virus H5N1. Front Vet Sci. 2023;10:
1286758 . - Skarlupka AL, Zhang X, Blas-Machado U, Sumner SF, Ross TM. Multi-influenza HA subtype protection of ferrets vaccinated with an N1 COBRA-based neuraminidase. Viruses. 2023;15:184.
- Uno N, Ebensen T, Guzman CA, Ross TM. Intranasal administration of octavalent next-generation influenza vaccine elicits protective immune responses against seasonal and pre-pandemic viruses. J Virol. 2024;98:
e0035424 . - Furey C, Scher G, Ye N, Kercher L, DeBeauchamp J, Crumpton JC, et al. Development of a nucleoside-modified mRNA vaccine against clade 2.3.4.4b H5 highly pathogenic avian influenza virus. Nat Commun. 2024;15:4350.
Figures
Table
Cite This ArticleOriginal Publication Date: February 21, 2025
Table of Contents – Volume 31, Number 3—March 2025
| EID Search Options |
|---|
|
|
|
|
|
|
Please use the form below to submit correspondence to the authors or contact them at the following address:
Valerie Le Sage, University of Pittsburgh, 3501 Fifth Ave, Pittsburgh, PA 15261, USA
Top